8.6: Вестибулярна система
- Page ID
- 88835

Медичний коледж Бейлора
Вестибулярна система функціонує для виявлення руху голови та положення щодо сили тяжіння і в першу чергу бере участь у тонкому контролі зорового погляду, постави, ортостазу, просторової орієнтації та навігації. Вестибулярні сигнали високо обробляються в багатьох областях мозку і беруть участь у багатьох найважливіших функціях. У цьому модулі ми надаємо огляд того, як працює вестибулярна система і як вестибулярні сигнали використовуються для керівництва поведінкою.
цілі навчання
- Визначте основні структури вестибулярної рецепторної системи.
- Опишіть нейроанатомію вестибулоокулярних, вестибулоспінальних та вестибуло-таламо-коркових шляхів.
- Охарактеризуйте вестибулярну комісуральну систему.
- Опишіть різні мультисенсорні області корки для сприйняття руху.
Вступ
Пам'ятайте запаморочення почуття, яке ви отримали в дитинстві після того, як ви стрибнули з каруселі або крутилися навколо, як вершина? Ці відчуття виникають внаслідок активації вестибулярної системи, яка виявляє наші рухи через простір, але не є свідомим почуттям, як зір чи слух. Насправді більшість вестибулярних функцій непомітні, але вестибулярні відчуття, такі як заколисування, можуть швидко спливати при їзді на американських гірках, вибоїстій їзді на літаку або плавання човна в бурхливих морях. Однак ці відчуття дійсно є побічними ефектами, і вестибулярна система насправді надзвичайно важлива для повсякденної діяльності, при цьому вестибулярні сигнали беруть участь у більшій частині обробки інформації мозку, яка контролює такі основні функції, як баланс, постава, стабілізація погляду, просторовий орієнтації та навігації, щоб назвати декілька. У багатьох областях мозку вестибулярна інформація поєднується з сигналами інших органів чуття, а також з руховою інформацією, що породжує сприйняття руху, усвідомлення тіла та поведінковий контроль. Тут ми вивчимо роботу вестибулярної системи та розглянемо деякі інтегровані обчислення, які мозок виконує за допомогою вестибулярних сигналів, щоб керувати нашою спільною поведінкою.

Будова вестибулярних рецепторів
Вестибулярні рецептори лежать у внутрішньому вусі поруч зі слуховою равликою. Вони виявляють обертальний рух (повороти голови), лінійний рух (переклади) та нахили голови щодо сили тяжіння і перетворюють ці рухи в нейронні сигнали, які можуть бути відправлені в мозок. У кожному вусі є п'ять вестибулярних рецепторів (модуль слуху, рис. 8.6.1- http://noba.to/jry3cu78), включаючи три напівкруглих каналу (горизонтальний, передній і задній), які транслюють обертальні кутові прискорення і два отолітових рецептора (шлуночка і saccule), які транслюють лінійні прискорення (Ліндеман, 1969). Разом напівкруглі канали та органи отоліту можуть реагувати на рух голови і підтримувати статичне положення голови щодо сили тяжіння у всіх напрямках у тривимірному просторі.
Ці рецептори містяться в серії з'єднаних між собою заповнених рідиною трубок, які захищені щільною, верхньою кісткою (Iurato, 1967). Кожен з трьох напівкруглих каналів лежить в площині, яка ортогональна до двох інших. Горизонтальний напівкруглий канал лежить в приблизно горизонтальній площині голови, тоді як передній і задній напівкруглі канали лежать вертикально в голові (Blanks, Curthoys, Bennett, & Markham, 1985). Напівкруглі рецепторні клітини каналу, які називаються волосяними клітинами, розташовані лише посередині круглих труб в спеціальному епітелії, покритому драглистою оболонкою, яка розтягується поперек трубки, утворюючи герметичне ущільнення, подібне до шкіри барабана (рис. 1А і 1В). Волосяні клітини так названі завдяки масиву майже 100 стереоцилій у шаховому порядку (як церковний орган труби), які виступають з верхньої частини клітини в вищерозташовану желатинову мембрану (Wersäll, 1956). Найкоротші стереоцилії знаходяться на одному кінці клітини, а найвищі на іншому (Ліндеман, 1969). При обертанні голови рідина в напівкруглих каналах відстає від руху голови і штовхає на желатинову мембрану, яка згинає стереоцилії.

Як показано на малюнку 8.6.2, коли голова рухається до рецепторних клітин волосся (наприклад, ліва голова повертається за лівий горизонтальний напівкруглий канал), стереоцилії згинаються до найвищого кінця і відкриваються спеціальні механічно закриті іонні канали в кінчиках вій, що збуджує ( деполяризує) клітину (Шотвелл, Джейкобс і Хадспет, 1981). Рух голови в протилежному напрямку викликає вигин в сторону найменших стереоцилій, що закриває канали і гальмує (гіперполяризує) клітину. Напівкруглі канали лівого і правого вуха мають протилежну полярність, тому, наприклад, при повороті голови вліво рецептори в лівому горизонтальному напівкруглому каналі будуть збуджуватися, а рецептори горизонтального каналу правого вуха будуть гальмуватися (рис. 8.6.3). Таке ж співвідношення справедливо і для вертикальних напівкруглих каналів. Вестибулярні аферентні нервові волокна іннервують основу волосяної клітини і збільшують або зменшують їх нервову швидкість стрільби, оскільки рецепторна клітина збуджується або гальмується (Dickman and Correia, 1989) відповідно, а потім несуть ці сигнали щодо обертального руху голови до мозку як частина вестибулокохлеарний нерв (Черепний нерв VIII). Вони потрапляють у стовбур мозку і закінчуються іпсилатеральними вестибулярними ядрами, мозочком та ретикулярною формацією (Carleton & Carpenter, 1984; Dickman & Fang, 1996). Первинною вестибулярною волосяною клітиною і аферентними нейромедіаторами є глутамат і аспартат. Завдяки механічним властивостям системи вестибулярних рецепторів, прискорення обертання голови інтегруються в сигнали швидкості (Van Egmond, Groen, & Jongkess, 1949), які потім кодуються аферентами напівкруглих каналів (Fernandez & Goldberg, 1971). Пороги виявлення обертального руху показали, що афференти можуть розрізняти різницю в швидкості голови близько 2 град/сек, але також чутливі до широкого діапазону природних рухів голови до високих швидкостей голови в сотнях град/сек (як ви могли б відчувати, коли ви робите швидкий поворот голови до гучного звуку, або виконують гімнастику; Sadeghi, Chacron, Taylor, & Cullen, 2007; Ю, Dickman, & Angelaki, 2012).
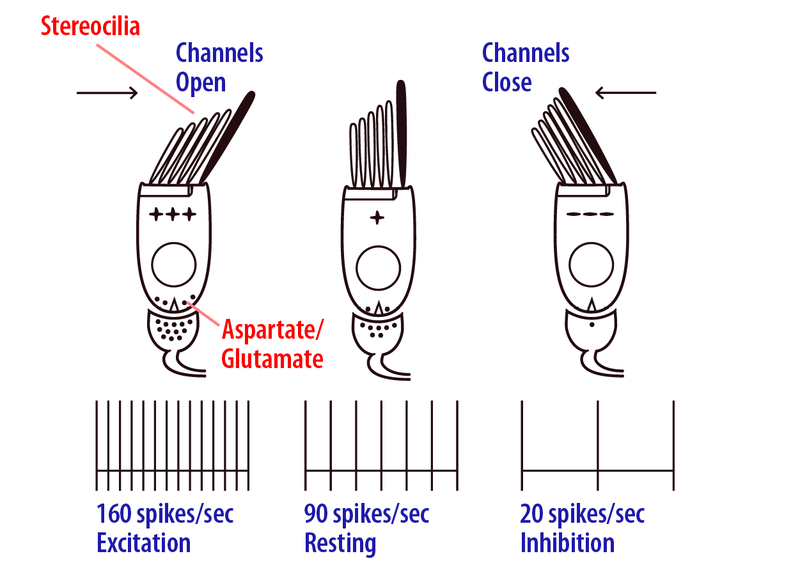

Отолітові рецептори чутливі до лінійних прискорень і нахилів голови щодо сили тяжіння (Fernandez & Goldberg, 1976a). Отолітовий рецептор шлуночки лежить паралельно горизонтальному напівкруглому каналу, а рецептор мішечки лежить вертикально в голові (модуль слуху, рис. 8.6.1- http://noba.to/jry3cu78). Як показано на малюнку 8.6.4, спеціальний отолітний епітелій містить рецепторні клітини волосся, стереоцилії яких поширюються на желатинову мембрану, яка покрита шаром кристалів карбонату кальцію, що називаються отоконієм, як породи, складені з утворенням пристані (Lindeman, 1969). Отоконії не впливають рухи рідини, але замість цього зміщуються лінійними прискореннями, включаючи переклади (наприклад, рухи вперед/назад або вгору/вниз) або зміни положення голови щодо сили тяжіння. Ці лінійні прискорення виробляють переміщення отоконію (через їх високу масу), подібно до каменів, що скочуються з пагорба, або ваша кавова чашка падає з приладової панелі автомобіля, коли ви натискаєте педаль газу. Рухи отокония згинають стереоцилію волосяної клітини і відкривати/закривають канали аналогічно тому, що описано для напівкруглих каналів. Однак отолітові волосяні клітини поляризовані таким чином, що найвищі стереоцилії спрямовані до центру сечовини і подалі від центру в мішечку, що ефективно розщеплює рецептори на дві протилежні групи (Flock, 1964; Lindeman, 1969). Таким чином, деякі клітини волосся збуджуються, а деякі гальмуються для кожної лінійної сили руху або нахилу голови, при цьому популяція рецепторів та їх іннервуючих аферентів спрямовано налаштована на всі рухи або нахили голови в 3D-просторі (Fernandez & Goldberg, 1976b).

Всі вестибулярні клітини волосся і афференти отримують з'єднання від вестибулярних ефектів, які представляють собою волокна, що проектують з мозку на органи вестибулярних рецепторів, функція яких недостатньо вивчена. Вважається, що ефекти контролюють чутливість рецептора (Boyle, Carey, & Highstein, 1991). Первинними ефектами нейромедіатора є ацетилхолін (Anniko & Arnold, 1991).
вестибулярні ядра
Вестибулярні ядра містять великий набір нервових елементів у стовбурі мозку, які отримують рух та інші мультисенсорні сигнали, потім регулюють реакції руху та сенсорний досвід. Багато нейрони вестибулярних ядер мають зворотні зв'язки з мозочком, які утворюють важливі регуляторні механізми контролю рухів очей, рухів голови та постави. Є чотири основні вестибулярні ядра, які лежать в ростральному мозку і хвостових понтах стовбура мозку; всі отримують прямий вхід від вестибулярних аферентів (Brodal, 1984; Precht & Shimazu, 1965). Багато з цих нейронів ядер отримують інформацію про конвергентний рух з протилежного вуха через інгібуючий комісуральний шлях, який використовує гамма-аміномасляну кислоту (ГАМК) як нейромедіатор (Kasahara & Uchino, 1974; Shimazu & Precht, 1966). Комісуральний шлях високоорганізований таким чином, що клітини, які отримують горизонтальні сигнали збуджуючого каналу від іпсилатерального вуха, також отримуватимуть контралатеральні гальмівні сигнали горизонтального каналу від протилежного вуха. Цей факт породжує «push-pull» вестибулярну функцію, завдяки якій спрямована чутливість до рух голови кодується протилежними рецепторними сигналами. Оскільки нейрони вестибулярних ядер отримують інформацію від двосторонніх рецепторів внутрішнього вуха і тому, що вони підтримують високу швидкість спонтанного стрільби (майже 100 імпульсів/сек), вони, як вважають, діють, щоб «порівняти» відносні швидкості розряду лівого проти правого каналу аферентної стрільби активності. Наприклад, під час повороту голови лівого стовбура мозку нейрони лівого стовбура мозку отримують інформацію про високу швидкість стрільби з лівого горизонтального каналу і інформацію про низьку швидкість стрільби з правого горизонтального каналу. Порівняння активності трактується як лівий поворот голови. Подібні реакції нейронів ядер існують, коли головка нахилена або згорнута, при цьому вертикальні напівкруглі канали стимулюються обертальним рухом у площині їх чутливості. Однак протилежний push-pull відповідь від вертикальних каналів відбувається з переднім напівкруглим каналом в одному вусі і суплощинним заднім напівкруглим каналом протилежного вуха. Пошкодження або захворювання, яке перериває інформацію про сигнал внутрішнього вуха з одного боку голови, може змінити нормальну активність спокою в VIIIth нервових аферентних волокнам і буде інтерпретуватися мозком як обертання голови, навіть якщо голова нерухома. Ці ефекти часто призводять до ілюзій спінінгу або обертання, що може бути досить засмучує і може спричинити нудоту або блювоту. Однак з часом комісуральні волокна забезпечують вестибулярну компенсацію - процес, за допомогою якого втрата односторонньої функції вестибулярних рецепторів частково відновлюється централізовано та поведінкові реакції, такі як вестибулоокулярний рефлекс (VOR) та постуральні реакції, переважно відновити (Беранек та ін., 2003; Феттер і Зе, 1988,; Ньюлендс, Гессен, Хак, & Анжелакі, 2001; Ньюлендс і Перахіо, 1990).
Окрім комісурального шляху, багато нейронів вестибулярних ядер отримують проприоцептивні сигнали від спинного мозку щодо руху та положення м'язів, зорові сигнали щодо просторового руху, інші мультисенсорні (наприклад, трійчасті) сигнали та сигнали вищого порядку від кори. Вважається, що коркові входи регулюють тонкий погляд і постуральний контроль, а також пригнічують нормальні компенсаторні рефлекси під час руху, щоб викликати вольові рухи. Особливе значення мають збігаються сигнали з напівкруглого каналу і отолітових аферентів, які дозволяють центральним вестибулярним нейронам обчислити специфічні властивості руху голови (Dickman & Angelaki, 2002). Наприклад, Ейнштейн (1907) показав, що лінійні прискорення рівнозначні незалежно від того, виникають вони від поступального руху або від нахилів голови щодо сили тяжіння. Рецептори отоліту не можуть розрізняти ці два, так як це, що ми можемо визначити різницю між тим, коли ми переводимо вперед і нахил назад, де лінійне прискорення, сигналізоване аферентами отоліту, однакове? Вестибулярні ядра та нейрони мозочка використовують конвергентні сигнали як від напівкруглих каналів, так і від рецепторів отоліту для розрізнення нахилу та перекладу, і, як наслідок, деякі клітини кодують нахил голови (Zhou, 2006), тоді як інші клітини кодують поступальний рух (Angelaki, Shaikh, Green, & Dickman, 2004).
Вестибулоокулярна система
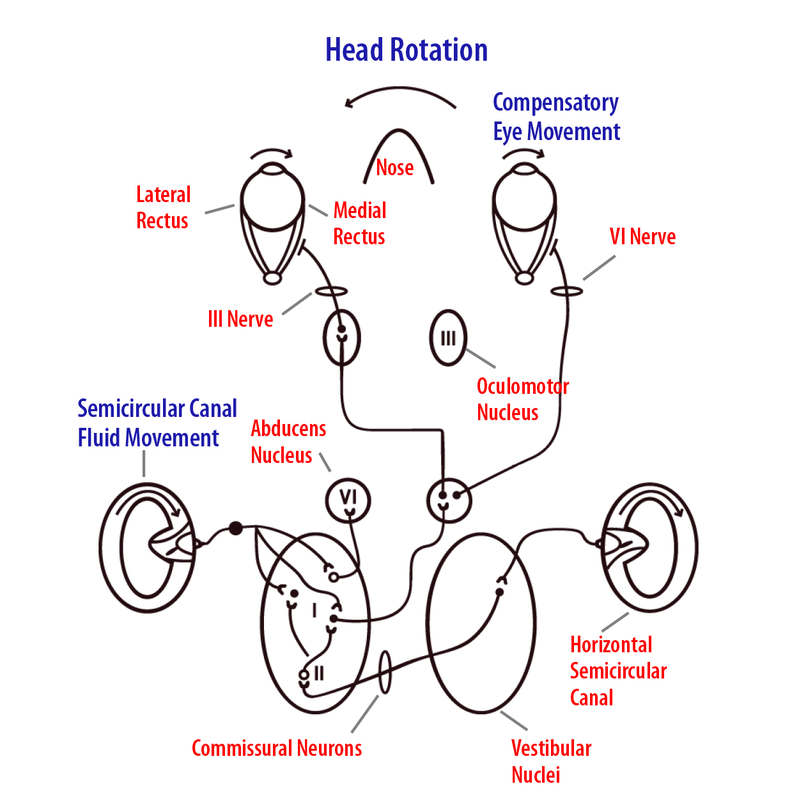
Вестибулярна система відповідає за контроль стійкості погляду під час руху (Crane & Demer, 1997). Наприклад, якщо ми хочемо читати знак у вітрині магазину під час прогулянки, ми повинні підтримувати фовеальну фіксацію на словах, компенсуючи комбіновані обертальні та поступальні рухи голови, що виникають під час нашого кроку. Вестибулярна система регулює компенсаторні рухи очей, шиї, хребта та кінцівок для підтримки погляду (Keshner & Peterson, 1995). Одним з основних компонентів, що сприяють стабільності погляду, є VOR, який виробляє рефлексивні рухи очей, рівні за величиною і протилежні напрямку до сприйнятого руху голови в 3D-просторі (Wilson et al., 1995). VOR настільки точний і швидкий, що дозволяє людям підтримувати візуальну фіксацію на об'єктах, що цікавлять, відчуваючи при цьому складні умови руху, такі як біг, катання на лижах, гра в теніс та водіння. Насправді стабілізація погляду у людей була показана повністю компенсаторною (по суті ідеальною) для більшості природної поведінки. Для отримання VOR вестибулярні нейрони повинні контролювати кожну з шести пар очних м'язів в унісон через певний набір з'єднань з окоруховими ядрами (Ezure & Graf, 1984). Передній і задній напівкруглі канали разом з мішечками контролюють вертикальні і крутильні (поворот ока навколо прямої видимості) рухи очей, в той час як горизонтальні канали і уриця контролюють горизонтальні рухи очей.
Щоб зрозуміти, як працює VOR, візьмемо приклад компенсаторного відгуку при повороті голови вліво під час читання слів на екрані комп'ютера. Основний шлях складається з горизонтальних напівкруглих каналів афферентів, які проектують на конкретні нейрони в вестибулярних ядрах. Ці ядра клітини, в свою чергу, посилають збудливий сигнал на контралатеральное відводить ядро, яке проектує через шостий черепний нерв для іннервування латерального прямого м'яза (рис. 8.6.5). Деякі викрадені нейрони відправляють збудливу проекцію назад через середню лінію до підрозділу клітин в іпсилатеральному окоруховому ядрі, яке, в свою чергу, проектує через третій черепний нерв для іннервування правої (іпсилатеральної) медіальної прямої м'язи. Коли робиться поворот голови вліво, вестибулярні аференти лівого горизонтального каналу збільшать швидкість стрільби і, отже, збільшать активність нейронів вестибулярних ядер, що проектують на протилежне (контралатеральне) праве відводить ядро. Викрадені нейрони виробляють скорочення правої бічної прямої кишки і через окрему проекцію клітини до лівого окорухового ядра збуджують ліві медіальні прямі м'язи. Крім того, відповідні двосторонні гальмівні з'єднання розслабляють ліву латеральну пряму і праву медіальну пряму очні м'язи. Отриманий рух правого ока для обох очей стабілізує об'єкт інтересу на сітківці для найбільшої гостроти зору.
Під час лінійних перекладів також зустрічається інший тип VOR (Paige & Tomko, 1991). Наприклад, рух боком вліво призводить до горизонтального руху очей вправо для підтримки зорової стійкості на об'єкті, що цікавить. Подібним чином вертикальні рухи голови вгору-вниз (наприклад, відбуваються під час ходьби або бігу) викликають протилежно спрямовані вертикальні рухи очей (Angelaki, McHenry, & Hess, 2000). Для цих рефлексів амплітуда поступального VOR залежить від відстані огляду. Це пов'язано з тим, що кут вергенції (тобто кут між лініями зору для кожного ока) змінюється в залежності від зворотної відстані до видимого зорового об'єкта (Schwarz, Busettini, & Miles, 1989). Зорові об'єкти, які знаходяться далеко (2 метри і більше), не вимагають кута розбіжності, але в міру наближення зорових об'єктів (наприклад, коли палець тримається близько до носа), потрібен великий кут нахилу. Під час поступального руху очі змінюватимуть кут нахилу, коли зоровий об'єкт рухається від близького до далі (або навпаки). Ці відповіді є результатом активації рецепторів отоліту, при цьому зв'язки з окоруховими ядрами аналогічні описаним вище для обертального вестибулоокулярного рефлексу. При нахилах голови отриманий рух очей називається крутінням, і складається з обертального руху очей навколо прямої видимості, яка знаходиться в напрямку, протилежному нахилу голови. Як вже говорилося вище, існують основні реципрокні зв'язки між вестибулярними ядрами і мозочком. Добре встановлено, що ці зв'язки мають вирішальне значення для адаптивного рухового навчання в вестибулоокулярному рефлексі (Lisberger, Pavelko, & Broussard, 1994).
Вестибуло-спинномозкова мережа

Існує два вестибулярних спадних шляху, які регулюють реакції м'язів тіла на рух і гравітацію, що складаються з латерального вестибуло-спинномозкового тракту (LVST) і медіального вестибуло-спинномозкового тракту (MVST). Рефлекторний контроль м'язів голови і шиї виникає через нейрони в медіальному вестибулоспінальному тракті (MVST). Ці нейрони містять швидкий вестибулоколічний рефлекс (VCR), який служить для стабілізації голови в просторі та бере участь у контролі погляду (Peterson, Goldber, Bilotto, & Fuller, 1985). Нейрони MVST отримують вхід від вестибулярних рецепторів і мозочка, а соматосенсорну інформацію зі спинного мозку. Нейрони MVST несуть як збудливі, так і гальмівні сигнали для іннервування згиначів шиї та розгиначів моторних нейронів у спинному мозку. Наприклад, якщо під час ходьби їздити по тріщині тротуару, нейрони MVST отримають низхідні та вперед сигнали лінійного прискорення від рецепторів отоліту та сигнали прискорення обертання вперед від вертикальних напівкруглих каналів. Відеомагнітофон буде компенсувати, надаючи збудливі сигнали спинним м'язам згиначів шиї та гальмівні сигнали до м'язів вентральних розгиначів шиї, які рухають голову вгору і навпаки падаючого руху, щоб захистити її від удару.
LVST включає топографічну організацію клітин вестибулярних ядер, які отримують значний вхід від мозочка, проприоцептивні входи зі спинного мозку та конвергентні аферентні сигнали від вестибулярних рецепторів. Волокна LVST проектують іпсилатеральні на багато рівнів рухових нейронів у шнурі, щоб забезпечити координацію різних груп м'язів для постурального контролю (Shinoda, Sugiuchi, Futami, Ando, & Kawasaki, 1994). Нейрони LVST містять ацетилхолін або глутамат як нейромедіатор і надають збудливий вплив на м'язові моторні нейрони розгиначів. Наприклад, волокна LVST виробляють розширення контралатеральної осьової мускулатури та мускулатури кінцівок при нахилі тіла вбік. Ці дії служать для стабілізації центру ваги тіла з метою збереження вертикальної постави.
Вестибуло-вегетативний контроль
Деякі нейрони вестибулярного ядра посилають проекції до ретикулярної формації, спинних понтинних ядер і ядра одиночного тракту. Ці з'єднання регулюють дихання та кровообіг за допомогою компенсаторних вестибулярних вегетативних реакцій, які стабілізують дихання та артеріальний тиск під час руху тіла та зміни щодо сили тяжіння. Вони також можуть бути важливими для індукції заколисування та блювоти.
Вестибулярні сигнали в таламусі і корі
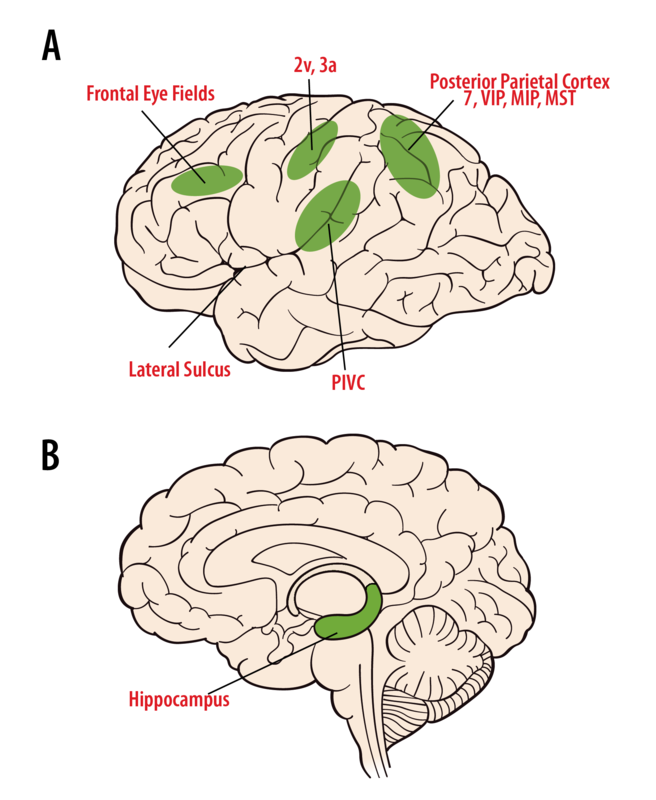
Когнітивне сприйняття руху, просторової орієнтації та навігації по простору виникає через мультисенсорну інформацію з вестибулярних, зорових і соматосенсорних сигналів в таламусі і корі (рис. 8.6.6). Нейрони вестибулярних ядер проектують двосторонньо на кілька таламічних областей. Нейрони в черевній задній групі реагують або на вестибулярні сигнали поодинці, або на вестибулярні плюс соматосенсорні сигнали, і проектує на первинну соматосенсорну кору (область 3а, 2v), соматосенсорну асоціацію кори, задню тім'яну кору (області 5 і 7), і ізолу скроневої кори (Marlinski & МакКрі, 2008; Менг, Травень, Дікман, & Ангелакі, 2007). Задня ядерна група (PO), поблизу медіального колінчастого тіла, отримує як вестибулярні, так і слухові сигнали, а також входи від верхнього колікула та спинного мозку, що вказує на інтеграцію множинних сенсорних сигналів. Деякі передні пульвінарні нейрони також реагують на подразники руху і проектують на коркову область 3a, задню інсулу та скронево-тім'яну кору (PIVC). У людини електростимуляція таламических областей виробляє відчуття руху і іноді запаморочення.
Клітини області 2v реагують на рух, і електрична стимуляція цієї області у людини виробляє відчуття руху, обертання або запаморочення. Область 3а лежить біля основи центральної борозни, що прилягає до рухової кори, і, як вважають, бере участь в інтегративному руховому управлінні головою та тілом (Guldin, Akbarian, & Grusser, 1992). Нейрони в PIVC є мультисенсорними, реагуючи на рух тіла, соматосенсорні, проприоцептивні та зорові стимули руху (Chen, DeAngelis, & Angelaki, 2011; Grusser, Pause, & Schreiter, 1982). PIVC і області 3а і 2v сильно пов'язані між собою. Вестибулярні нейрони також спостерігалися в задній тім'яній корі; в області 7, в вентральній внутрішньотім'яної області (VIP), медіальна внутрішньотім'яна область (MIP) та медіальна верхня скронева область (MST). VIP містить мультимодальні нейрони, що займаються просторовим кодуванням. Нейрони MIP та MST реагують на рух тіла через простір шляхом мультисенсорної інтеграції зорового руху та вестибулярних сигналів (Gu, DeAngelis, & Angelaki, 2007), і багато клітин MST безпосередньо беруть участь у сприйнятті заголовка (Gu, Watkins, Angelaki, & DeAngelis, 2006). Ураження тім'яних ділянок корки може призвести до плутанини в просторовій обізнаності. Нарешті, райони, пов'язані з контролем саккад та рухів очей переслідування, включаючи область 6, область 8 та верхню лобову звивину, отримують вестибулярні сигнали (Фукусіма, Сато, Фукусіма, Шінмей та Канеко, 2000). Як ці різні коркові області сприяють нашому сприйняттю руху та просторової орієнтації, досі недостатньо вивчено.
Просторова орієнтація і навігація
Наша здатність знати, де ми знаходимося, і орієнтуватися в різних просторових місцях є важливою для виживання. Вважається, що пізнавальна карта нашого середовища створюється шляхом дослідження, а потім використовується для просторової орієнтації та навігації, наприклад, їзди до магазину або прогулянки темним будинком (McNaughton, Battaglia, Jensen, Moser, & Moser, 2006). Клітини лімбічної системи та гіпокампу, які сприяють цим функціям, були ідентифіковані, включаючи клітини місця, клітини сітки та клітини напрямку голови (рис. 6B). Місце клітини в гіпокампі кодують конкретні місця в навколишньому середовищі (O'Keefe, 1976). Клітини сітки в енторинальній корі кодують просторові карти за тессельованим малюнком (Hafting, Fyhn, Molden, Moser, & Moser, 2005). Клітини напрямку голови в передньо-спинному таламусі кодують напрямок руху, незалежно від просторового розташування (Taube, 1995). Вважається, що ці типи клітин працюють разом, щоб забезпечити просторову орієнтацію, просторову пам'ять та нашу здатність орієнтуватися. Як клітини місця, так і клітини напрямку голови залежать від функціонуючої вестибулярної системи для підтримки їх спрямованості та орієнтації інформації (Stackman, Clark, & Taube, 2002). Шляхи, за допомогою яких вестибулярні сигнали потрапляють до навігаційної мережі, недостатньо зрозумілий; однак пошкодження вестибулярної системи, гіпокампу та спинного таламуса часто порушує нашу здатність орієнтуватися у звичних умовах, переміщатися з місця на місце або навіть знаходити дорогу додому.
Заколисування

Хоча ряд умов може спричинити заколисування, зазвичай вважається, що це викликано невідповідністю сенсорних сигналів між вестибулярними, зоровими та проприоцептивними сигналами (Yates, Miller, & Lucot, 1998). Наприклад, читання книги в автомобілі на звивистій дорозі може спричинити заколисування, внаслідок чого прискорення, які відчуває вестибулярна система, не відповідають зоровому вводу. Однак, якщо хтось дивиться у вікно на декорації, що проходять під час тієї ж подорожі, жодна хвороба не виникає, оскільки візуальні та вестибулярні сигнали знаходяться у вирівнюванні. Морська хвороба, форма заколисування, представляється особливим випадком і виникає внаслідок незвичайного вертикального коливального і рухового руху. Дослідження на людях виявили, що низькочастотні коливання 0,2 Гц і великі амплітуди (наприклад, виявлені у великих морях під час шторму), швидше за все, спричиняють заколисування, причому більш високі частоти пропонують невеликі проблеми.
Резюме
Тут ми бачили, що вестибулярна система перетворює та кодує сигнали про рух голови та положення щодо сили тяжіння, інформації, яка потім використовується мозком для багатьох важливих функцій та поведінки. Ми насправді багато розуміємо щодо вестибулярного внеску до фундаментальних рефлексів, таких як компенсаторні рухи очей та баланс під час руху. Більш пізній прогрес був досягнутий до розуміння того, як вестибулярні сигнали поєднуються з іншими сенсорними сигналами, такими як зір, в таламусі та корі, щоб викликати сприйняття руху. Однак існує багато складних когнітивних здібностей, які, як ми знаємо, вимагають вестибулярної інформації для функціонування, наприклад, просторової орієнтації та навігаційної поведінки, але ці системи лише починають досліджуватися. Майбутні дослідження щодо функції вестибулярної системи, ймовірно, будуть спрямовані на пошук відповідей на питання щодо того, як мозок справляється з втратою вестибулярного сигналу. Насправді, за даними Національного інституту охорони здоров'я, майже 35% американців у віці старше 40 років (69 мільйонів людей) повідомили про хронічні проблеми, пов'язані з вестибулярними захворюваннями. Тому для здоров'я людини важливо краще зрозуміти, як вестибулярні сигнали сприяють загальним функціям мозку та як можна реалізувати кращі варіанти лікування вестибулярної дисфункції.
Зовнішні ресурси
- Анімоване відео вестибулярної системи
- http://sites.sinauer.com/neuroscienc...ions14.01.html
Питання для обговорення
- Якщо людина підтримує втрату вестибулярних рецепторів в одному вусі через хворобу або травму, якими симптомами страждала б людина? Чи будуть симптоми постійними?
- Часто заколисування полегшується, коли людина дивиться на далекі предмети, такі як речі, розташовані на дальньому горизонті. Чому перегляд на далекій відстані допомагає при хворобі руху, тоді як перегляд на близькій відстані (наприклад, читання карти чи книги) погіршує ситуацію?
- Вестибулярні сигнали поєднуються з зоровими сигналами в певних областях кори і допомагають у сприйнятті руху. Які типи сигналів забезпечує зорова система для самостійного руху через простір? Які типи вестибулярних сигналів узгоджувалися б з обертальним та поступальним рухом?
Лексика
- Викрадає ядро
- Група збудливих рухових нейронів в медіальному стовбурі мозку, які посилають проекції через VI-й черепний нерв для контролю іпсилатеральної латеральної прямої м'язи. Крім того, викрадені інтернейрони посилають збудливу проекцію по середній лінії до підрозділу клітин в іпсилатеральному окоруховому ядрі, які проектують через III черепний нерв для іннервування іпсилатеральної медіальної прямої м'язи.
- ацетилхолін
- Органічна сполука нейромедіатор, що складається з оцтової кислоти і холіну. Залежно від типу рецептора, ацетихолін може мати збудливу, інгібуючу або модулюючу дію.
- Аферентні нервові волокна
- Поодинокі нейрони, які іннервують рецепторні волосяні клітини і несуть вестибулярні сигнали до мозку як частина вестибулокохлеарного нерва (черепний нерв VIII).
- Аспартат
- Збуджуючий амінокислотний нейромедіатор, який широко використовується вестибулярними рецепторами, аферентами та багатьма нейронами мозку.
- компенсаторні рефлекси
- Стабілізуючий моторний рефлекс, який виникає у відповідь на сприйнятий рух, наприклад вестибулоокулярний рефлекс, або постуральні реакції, що виникають під час бігу або катання на лижах.
- Деполяризований
- Коли рецепторні клітини волосся мають механічно закриті канали, клітина збільшує свою мембранну напругу, що виробляє вивільнення нейромедіатора для збудження іннервируючого нервового волокна.
- Пороги виявлення
- Найменша кількість руху голови, про яку може достовірно повідомити спостерігач.
- спрямований тюнінг
- Бажаний напрямок руху, який демонструють клітини волосся та афференти там, де відбувається пікова збудлива реакція та найменш бажаний напрямок, де не відбувається реакції. Кажуть, що клітини «налаштовані» на найкращий і найгірший напрямок руху, причому між напрямками руху викликають меншу, але спостережувану реакцію.
- Гамма-аміномасляна кислота
- Основний гальмівний нейромедіатор у вестибулярній комісуральній системі.
- Стабільність погляду
- Поєднання реакцій очей, шиї та голови, які узгоджені для підтримки зорової фіксації (fovea) на цікаву точку.
- глутамат
- Збуджуючий амінокислотний нейромедіатор, який широко використовується вестибулярними рецепторами, аферентами та багатьма нейронами мозку.
- Волосяні клітини
- Рецепторні клітини вестибулярної системи. Вони називаються волосяними клітинами через безліч волосяних війок, які простягаються від верхівкової поверхні клітини в желатинову мембрану. Механічні закриті іонні канали в кінчиках вій відкриваються і закриваються, коли вії згинаються, щоб викликати зміни напруги мембрани у волосяній клітині, пропорційні інтенсивності та напрямку руху.
- гіперполяризує
- Коли рецепторні клітини волосся мають механічно закриті канали, клітина зменшує свою мембранну напругу, що виробляє менший вивільнення нейромедіаторів для пригнічення іннервуючого нервового волокна.
- Латеральна пряма м'яз
- Очний м'яз, що повертається назовні в горизонтальній площині.
- Бічні вестибуло-спинномозкові шляхи
- Вестибулярні нейрони, які проектують на всі рівні спинного мозку на іпсилатеральній стороні для контролю постави та балансування рухів.
- Механічно закриті іонні канали
- Іонні канали, розташовані в кінчиках стереоцилій на рецепторних клітинам, які відкриваються/закриваються, коли вії згинаються відповідно до найвищих/найменших війок. Ці канали проникні для іонів калію, яких багато в рідині, що купає верхню частину волосяних клітин.
- Медіальний вестибуло-спинномозковий тракт
- Нейрони вестибулярного ядра двосторонньо проектують на шийні моторні нейрони хребта для контролю руху голови та шиї. Тракт головним чином функціонує у напрямку погляду та стійкості під час руху.
- Нейромедіатори
- Хімічна сполука, яка використовується для відправки сигналів від рецепторної клітини до нейрона, або від одного нейрона до іншого. Нейромедіатори можуть бути збудливими, інгібуючими або модулюючими і упаковані в невеликі бульбашки, які звільняються з кінцевих терміналів клітин.
- окорухові ядра
- Включає в себе три нейрональні групи в стовбурі мозку, відхідне ядро, окорухове ядро і трохлеарне ядро, клітини якого посилають рухові команди шести парам очних м'язів.
- Окорухове ядро
- Група клітин середнього стовбура мозку, які містять підгрупи нейронів, які проектують на медіальну пряму кишку, нижню косу, нижню пряму кишку та верхні прямі м'язи очей через 3-й черепний нерв.
- Отоконія
- Дрібні частинки карбонату кальцію, які упаковані в шар поверх желатинної мембрани, яка покриває стереоцилії клітин волосся отолітового рецептора.
- Отолітові рецептори
- Два вестибулярних рецептора внутрішнього вуха (уриця і мішечок), які перетворюють лінійні прискорення і нахил голови щодо сили тяжіння в нервові сигнали, які потім передаються в мозок.
- Пропріоцептивний
- Сенсорна інформація щодо положення та руху м'язів, що виникають з рецепторів у м'язах, сухожиллям та суглобах.
- Напівкруглі канали
- Набір з трьох внутрішніх вушних вестибулярних рецепторів (горизонтальних, передніх, задніх), які перетворюють прискорення обертання голови в головні сигнали швидкості обертання, які потім передаються в мозок. У кожному вусі є три напівкруглих каналу, причому основні площини кожного каналу ортогональні один до одного.
- Стереоцилія
- Волосяні виступи з верхньої частини рецепторних клітин волосся. Стереоцилії розташовані по висхідній висоті і при зміщенні в бік найвищих війок механічні закриті канали відкриваються і клітина збуджується (деполяризується). Коли стереоцилії зміщуються в бік найдрібніших війок, канали закриваються і клітина гальмується (гіперполяризується).
- Торсіон
- Обертальний рух очей навколо прямої видимості, яка складається з напрямку за годинниковою стрілкою або проти годинникової стрілки.
- Кут вергенції
- Кут між лінією зору для двох очей. Низькі кути вергенції вказують на далекоглядні об'єкти, тоді як великі кути вказують на огляд ближніх об'єктів.
- Вестибулярна компенсація
- Після травмування однієї сторони вестибулярних рецепторів або вестибулокохлеарного нерва нейрони центральних вестибулярних ядер поступово відновлюють більшу частину своєї функції за допомогою механізмів пластичності. Однак відновлення ніколи не буває повним, а екстремальні середовища руху можуть призвести до запаморочення, нудоти, проблем з рівновагою та просторовою пам'яттю.
- вестибулярні ефекти
- Нервові волокна, що походять від ядра в стовбурі мозку, які проектують від мозку, щоб іннервувати волосяні клітини вестибулярних рецепторів та аферентні нервові термінали. Ефференти мають модулюючу роль на своїх цілях, яка недостатньо зрозуміла.
- Вестибулярна система
- Складається з набору рецепторів виявлення руху та сили тяжіння у внутрішньому вусі, набору первинних ядер у стовбурі мозку та мережі шляхів, що несуть сигнали руху та гравітації до багатьох областей мозку.
- Вестибулокохлеарний нерв
- VIII черепний нерв, який несе волокна, що іннервують вестибулярні рецептори і равлику.
- вестибулоокулярний рефлекс
- Рухи очей, вироблені вестибулярним стовбуром мозку, рівні за величиною і протилежні в напрямку руху голови. VOR функціонує для підтримки візуальної стабільності в цікавій точці і майже ідеально підходить для всіх природних рухів голови.
Посилання
- Анжелакі, Д.Е., МакГенрі, М.К., & Гесс, Б.Дж. (2000). Приматні поступальні вестибулоокулярні рефлекси. Високочастотна динаміка і тривимірні властивості при бічному русі. Нейрофізіол 83, 1637—1647.
- Анжелакі, Д. Е., Шейх, А.Г., Грін, А.М., & Дікман, Дж. Нейрони обчислюють внутрішні моделі фізичних законів руху. Природа, 430, 560—564.
- Анніко, М., Арнольд, В. (1991). Локалізація рецепторів ацетилхоліну у дорослих кохлеарних і вестибулярних клітинам волосся людини. АС і отолар, 111, 491-499.
- Беранек, М., Хашемауї, М., Іду, Е., Ріс, Л., Уно, А., Годо, Е. Віберт, Н. Тривала пластичність іпсилезіональних нейронів медіального вестибулярного ядра після односторонньої лабіринтектомії. J Нейрофізіол, 90, 184—203.
- Бланкс, Р.Г.І., Кертойс, І.С., Беннетт, М.Л., і Маркхем, К.Х. (1985). Площинні зв'язки напівкруглих каналів у резус-мавп і білок. Мозок Рез, 340, 315—324.
- Бойл, Р., Кері, Дж. П., і Хайштейн, С.М. (1991). Морфологічні кореляти динаміки реакції та еферентної стимуляції в горизонтальних напівкруглих каналах аферентів жаби, Opsanus tau. J Нейрофізіол, 66, 1504—1511.
- Бродаль, А. Вестибулярні ядра у мавпи макаки. Дж. комп. Неврол., 227, 252-266.
- Карлтон, С.К., і Карпентер, М.Б. (1984). Розподіл первинних вестибулярних волокон в стовбурі мозку і мозочку мавпи. Мозок Рез, 294, 281—298.
- Чен, А., ДеАнджеліс, Г.К., & Анжелакі, Д.Е. (2011). Порівняння вестибулярної просторово-темпоральної настройки в парієтоінсулярної вестибулярної корі макаки, вентральної внутрішньотім'яної області та медіальної верхньої скроневої області. Неврологія, 31, 3082—3094.
- Крейн, Б.Т., і Демер, Дж. Л. (1997). Стабілізація погляду людини під час природної діяльності: трансляції, обертання, збільшення та ефекти відстані цілі. J Нейрофізіол, 78, 2129-2144.
- Дікман, Дж., і Анжелакі, Д.Е. (2002). Вестибулярні схеми конвергенції в нейроні вестибулярних ядер оповіщення приматів. Нейрофізіолог., 88, 3518—3533.
- Дікман, Дж., і Коррея, М.Дж. (1989). Відгуки голуба горизонтальний напівкруглий канал аферентних волокон. I. ступінчаста, трапецієподібна і низькочастотна синусоїдна механічна і обертальна стимуляція. J Нейрофізіол, 62, 1090—1101.
- Дікман, Дж., & Ікла, Q. (1996). Диференціальні центральні проекції вестибулярних аферентів у голубів. J Комп Нейрол, 367, 110—121.
- Ейнштейн, А. (1907) .Про принцип відносності і висновки, зроблені з нього. Джарб. Радіостанція, 4, 41-462.
- Езуре, К., Граф К.А. Проведено кількісний аналіз просторової організації вестибуло-очних рефлексів у бічних і фронтальнооких тварин. Орієнтація напівкруглих каналів і екстраокулярних м'язів. Неврологія, 12, 85—94.
- Фернандес, К., і Голдберг, Дж. М. (1976b). Фізіологія периферичних нейронів, що іннервують отолітні органи мавпи білки. II. Спрямована вибірковість і відносини сили реагування. Нейрофізіол., 39, 985—995.
- Фернандес, К., і Голдберг, Дж. М. (1971). Фізіологія периферичних нейронів, що іннервують напівкруглі канали мавпи білки. II. Реакція на синусоїдальну стимуляцію і динаміку периферичної вестибулярної системи. J Нейрофізіол, 34, 661—675.
- Фернандес, К., і Голдберг, Дж. М. (1976а). Фізіологія периферичних нейронів, що іннервують отолітового органу у мавпи білки. Реакція на статичні нахили і на тривалу відцентрову силу. Нейрофізіол., 39, 970—984.
- Феттер, М., & Зі, Д.С. (1988). Відновлення після односторонньої лабіринтектомії у резус-мавпи. J Нейрофізіол, 59, 370—393.
- Флок, А. Структура макули з особливим посиланням на спрямовану взаємодію сенсорних реакцій, виявлену морфологічною поляризацією. J клітинний біол, 22, 413-431.
- Фукусіма, К., Сато, Т., Фукусіма, Шінмей, Ю., і Канеко С.Р. (2000). Активність плавного переслідування пов'язаних нейронів у мавпи пердугоподібних кортесах під час переслідування та пасивного обертання всього тіла. Нейрофіс., 83, 563—587.
- Груссер О., Пауза, М., Шрайтер, У. Нейронні реакції в тім'яно-інсулярної вестибулярної корі тривожних мавп Java (Macaca fascicularis). У А.Руку та М.Кроммелінка (ред.), Фізіологічні та патологічні аспекти рухів очей. Haque: W. небажані видавці.
- Гу, Ю., ДеАнджеліс, Г.К., & Анжелакі, Д.Е. (2007). Функціональний зв'язок між областю mstd і сприйняттям заголовка на основі вестибулярних сигналів. Нат Неврологія, 10, 1038—1047.
- Гу, Ю., Уоткінс, П.В., Анжелакі, Д. Е., & ДеАнджеліс, Г.К. (2006). Візуальний та невізуальний внесок у тривимірну селективність заголовків у медіальній верхній скроневій області. Дж. Неврологія, 26, 73—85.
- Гулдін О.О., Акбарян С.І., Груссер О.М. Кортико-коркові зв'язки та цитоархітектоніка вестибулярної кори приматів: дослідження у мавп білок (Saimiri sciureus). Дж. комп. Нейрол., 326, 375—401.
- Хафтінг, Т., Файн, М., Молден, С., Мозер, М.-Б., Мозер, Е.І. (2005). Мікроструктура просторової карти в енторинальній корі. Природа, 436, 801—806.
- Юрато С. Субмікроскопічна будова внутрішнього вуха. Лондон, Англія: Пергамон Прес.
- Касахара М., Учино Ю. Двосторонні напівкруглі вхідні канали до нейронів у вестибулярних ядрах кішки. Експ Мозок Рес, 20, 285—296.
- Кешнер, Е.А., і Петерсон, Б.В. (1995). Механізми, що контролюють стабілізацію голови людини. Динаміка голови і шиї при випадкових обертаннях в горизонтальній площині. J Нейрофізіол, 73, 2293-2301.
- Ліндеман, Г. Дослідження морфології сенсорних областей вестибулярного апарату з 45 цифрами. Ергеб Анат Ентвіклунггеш, 42, 1—113.
- Лісбергер С.Г., Павелко Т.А., Бруссар Д.М. (1994). Нейронні основи рухового навчання в вестибулоокулярному рефлексі приматів. Зміни реакцій стовбурових нейронів мозку. Нейрофізіол, 72, 928—973.
- Марлінський В.І., МакКрі Р.А. (2008). Активність вентрозадніх нейронів таламуса при обертанні і трансляції в горизонтальній площині у тривожної білки мавпи. J Нейрофізіол, 99, 2533-2545.
- Макнотон, Б.Л., Батталья, Ф.П., Дженсен, О., Мозер, Е.І., і Мозер, М.-Б. (2006). Інтеграція шляху та нейронна основа «когнітивної карти». Преподобний Природа, 7, 663—678.
- Менг, Х., Мей, П.Дж., Дікман, Дж., & Анжелакі, Д. Е. (2007). Вестибулярні сигнали у приматного таламуса: властивості та походження. Дж. Неврологія, 27, 13590—13602.
- Ньюлендс, С.Д., і Перачіо, А.А. (1990). Компенсація активності, пов'язаної з горизонтальним каналом в медіальному вестибулярному ядрі після односторонньої лабіринтної абляції в декребратної піщанці, тобто нейрони типу I. Експ Мозок Рес, 82, 359—372.
- Ньюлендс, С.Д., Гессен, С.В., Хак, А., і Анжелакі, Д. Е. (2001). Голова нестримного горизонтального погляду зміщується після односторонньої лабіринтектомії у резус-мавпи. Експ Мозок Рес, 140, 25—33.
- О'Кіф, Дж. (1976). Помістіть юнітів в гіпокампі вільно рухається щура. Експ. Нейрол, 51, 78—109.
- Пейдж, Г.Д., і Томко Д.Л. (1991). Рух очей реагує на лінійний рух голови у мавпи білки. Основні характеристики. J Нейрофізіол, 65, 1170—1182.
- Петерсон, Б.В., Голдберг, Дж., Білотто, Г., і Фуллер, Дж. Цервікоколічний рефлекс: його динамічні властивості та взаємодія з вестибулярними рефлексами. J Нейрофізіол, 54, 90—109.
- Прехт, В., і Шимазу, Х. (1965). Функціональні зв'язки тонічних і кінетичних вестибулярних нейронів з первинними вестибулярними афферентами. J Нейрофізіол, 28, 1014—1028.
- Садегі, С.Г., Чакрон, М.Дж., Тейлор, М.К., і Каллен, К.Е. (2007). Нейронна мінливість, пороги виявлення та передача інформації в вестибулярній системі. Неврологія, 27, 771—781.
- Шварц, У., Бусеттіні, К., & Майлз, Ф.А. (1989). Очні реакції на лінійний рух обернено пропорційні відстані перегляду. Наука, 245, 1394—1396.
- Шимадзу, Х., і Прехт, В. (1966). Пригнічення центральних вестибулярних нейронів з контралатерального лабіринту і його опосередковуючого шляху. J Нейрофізіол, 29, 467—492.
- Шинода, Ю., Суджучі, Ю., Футамі, Т., Андо, Н., & Кавасакі, Т. (1994). Вхідні шаблони і шляхи від шести напівкруглих каналів до мотонейронів м'язів шиї. I. Багатофункціональна група м'язів. Нейрофізіол., 72, 2691—2802.
- Шотвелл, С.Л., Джейкобс, Р., і Хадспет, А.Дж. (1981). Спрямована чутливість окремих клітин хребетних волосся до контрольованого відхилення їх волосяних пучків. АНЬЯСА, 374, 1—10.
- Стекман, Р.В., Кларк, А.С., і Таубе, Дж. (2002). Гіпокампальні просторові уявлення вимагають вестибулярного введення. Гіпокамп, 12, 291—303.
- Таубе Дж. С. Клітини напрямку голови реєструються в передніх таламічних ядрах вільно рухаються щурів. Невронаук, 15, 70—86.
- Ван Егмонд, А.А., Гроен, Дж., і Джонкесс, Л.Б. (1949). Механіка напівкруглих каналів. J Фізіолова навантаження, 110, 1—17.
- Версалл, Дж. (1956). Дослідження будови та іннервації сенсорного епітелію кристоподібних ампул у морської свинки; світлове та електронно-мікроскопічне дослідження. Акта Отоларингол Супп, 126, 1—85.
- Вілсон, В.Дж., Бойл, Р., Фукусіма, К., Роуз, П.К., Шинода, Ю., Суджучі, Ю., & Учіно, Ю. (1995). Вестибулокольний рефлекс. J Вестин Рес, 5, 147—170.
- Єйтс, Б.Дж., Міллер А.Д., і Люкот, Дж. Фізіологічні основи та фармакологія заколисування: оновлення. Мозок Рес Бик, 47, 395-406.
- Ю., Дж., Дікман, Дж., Анжелакі, Д.Е. (2012). Пороги виявлення афектів макаки отоліту. Дж. Невронаук, 32, 8306—8316.
- Чжоу, В., Тан, Б.Ф., Ньюлендс, С.Д., Кінг, WM (2006). Відповіді мавпячих вестибулярних нейронів на трансляцію і кутове обертання. J Нейрофізіол 96, 2915-2930.