4.2: Зір- Сітківка
- Page ID
- 72228
Анатомія сітківки
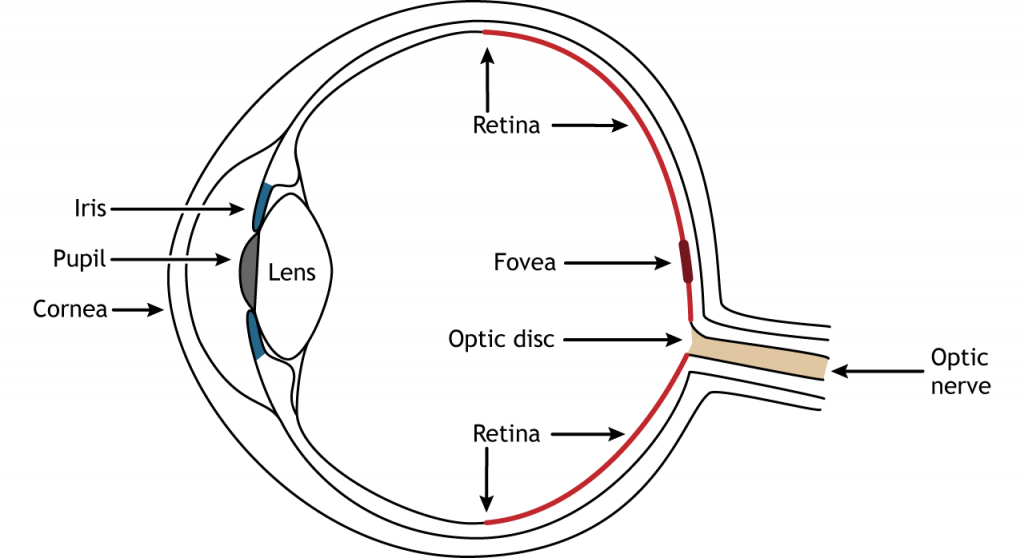
Передня частина ока складається з рогівки, зіниці, райдужної оболонки та кришталика. Рогівка - це прозора, зовнішня частина ока. Він охоплює зіницю і райдужну оболонку і є першим місцем заломлення світла. Зіниця - це отвір в райдужній оболонці, що дозволяє світлу потрапляти в око. Райдужна оболонка - це кольорова частина ока, яка оточує зіницю і разом з місцевими м'язами може контролювати розмір зіниці, щоб забезпечити відповідну кількість світла, щоб потрапити в око. Кришталик розташовується позаду зіниці і райдужної оболонки. Лінза заломлює світло, щоб сфокусувати зображення на сітківці. Правильне фокусування вимагає, щоб лінза розтягнулася або розслаблялася, процес називається акомодацією.
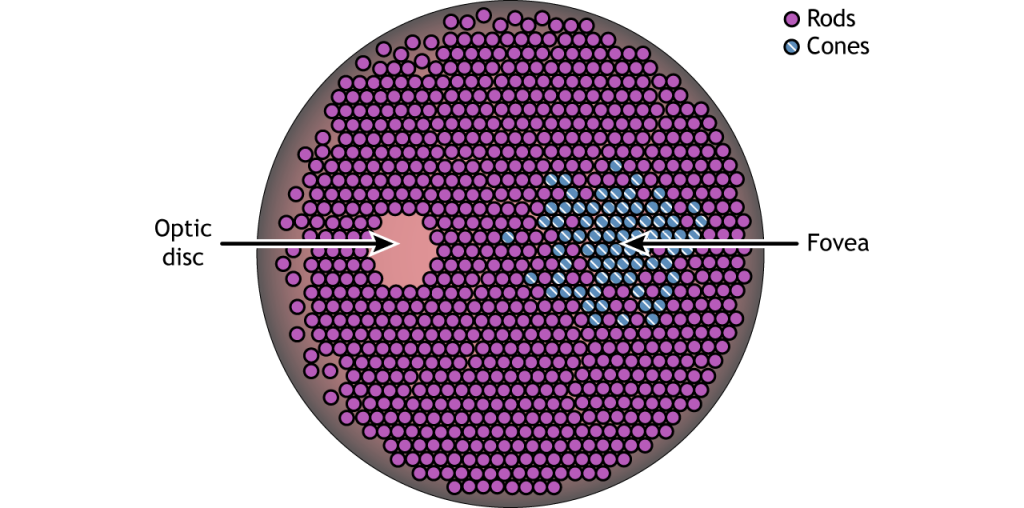
Сітківка - це світлочутлива область в задній частині ока, де розташовані фоторецептори, спеціалізовані клітини, які реагують на світло. Сітківка покриває всю задню частину ока, тому вона має форму миски. Посередині чаші знаходиться ямка, область найвищої гостроти зору, що означає область, яка може формувати найчіткіші зображення. Зоровий нерв проектує на мозок із задньої частини ока, несучи інформацію з клітин сітківки. Там, де йде зоровий нерв, немає фоторецепторів, оскільки аксони від нейронів збираються разом. Ця область називається диском зорового нерва і є місцем розташування сліпої плями в нашому полі зору.
Клітини сітківки
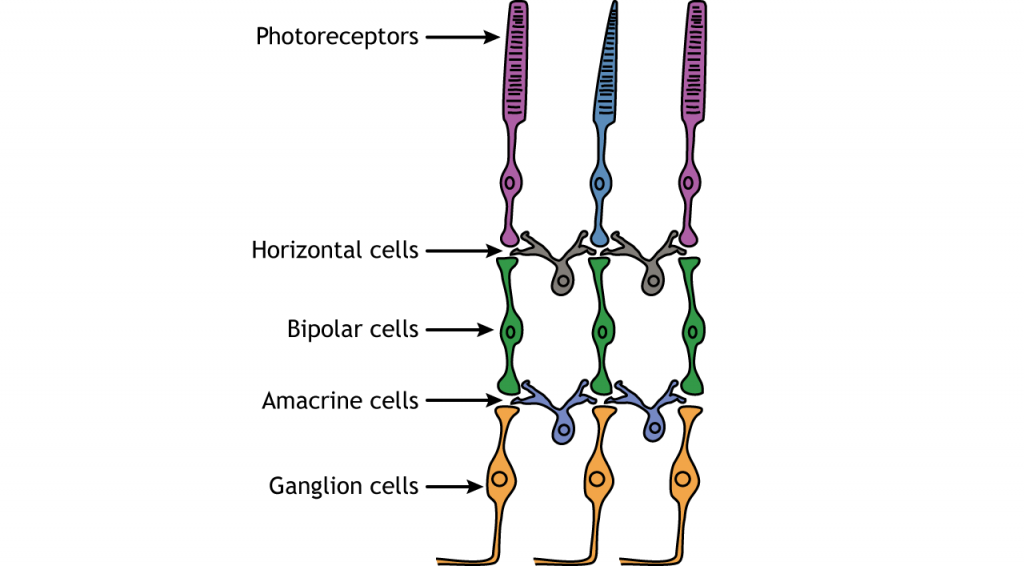
Крім фоторецепторів, в сітківці є ще чотири типи клітин. Фоторецептори синапсують на біполярних клітині, а біполярні клітини синапсують на гангліозних клітині. Горизонтальні і амакринні клітини дозволяють здійснювати зв'язок в латеральному напрямку між нейронами.
Напрямок інформації
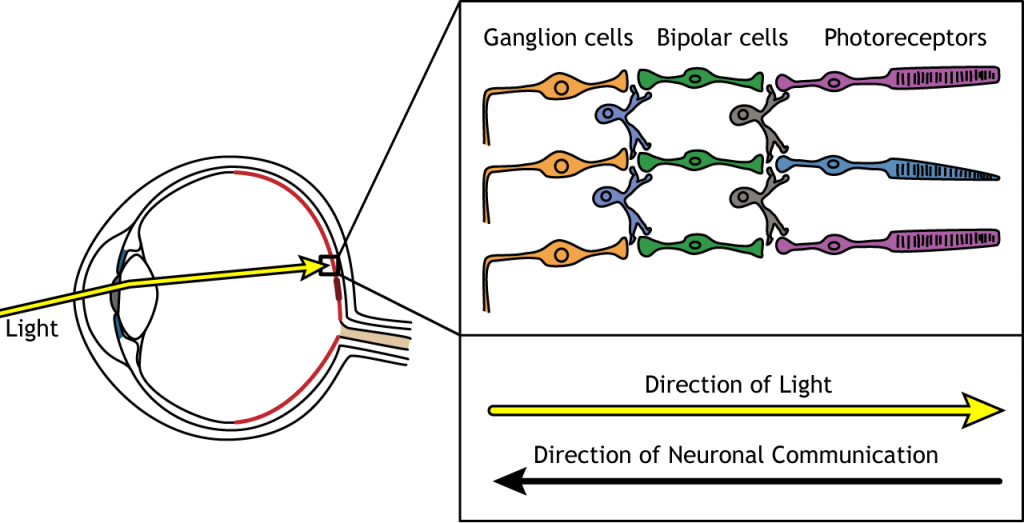
Коли світло потрапляє в око і вражає сітківку, він повинен пройти через всі шари нейрональних клітин, перш ніж досягти і активувати фоторецептори. Потім фоторецептори ініціюють синаптичну зв'язок назад до гангліозних клітин.
рецептори
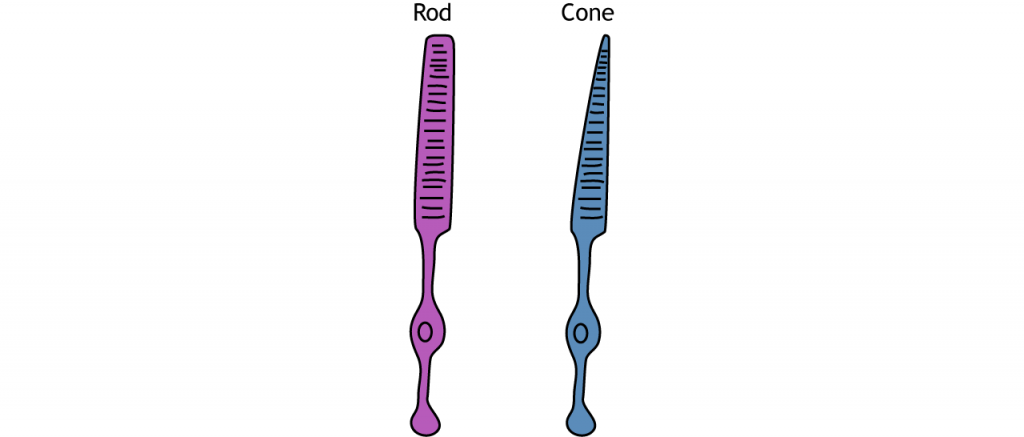
Фоторецептори - це спеціалізовані рецептори, які реагують на світло. Існує два типи фоторецепторів: палички і шишки. Стрижні більш чутливі до світла, що робить їх головним чином відповідальними за зір в умовах низької освітленості, як вночі. Шишки менш чутливі до світла і найбільш активні в умовах денного світла. Шишки також відповідають за кольоровий зір.
Щільність рецепторів
Окрім того, що мають різні зорові функції, стрижні та конуси також розподіляються по сітківці в різній щільності. Шишки в першу чергу виявляються в ямочці, області сітківки з найвищою гостротою зору. Решта сітківки - переважно стрижні. Область диска зорового нерва не має фоторецепторів, оскільки аксони гангліозних клітин виходять з сітківки і утворюють зоровий нерв.
Фототрансдукція
Фоторецептори відповідають за сенсорну трансдукцію в зоровій системі, перетворюючи світло в електричні сигнали в нейрони. Для наших цілей, щоб вивчити функцію фоторецепторів, ми будемо А) зосереджуватися на чорно-білому світлі (не кольоровий зір) і Б) припустити, що клітини рухаються або з області темряви в область світла, або навпаки.
Фоторецептори не розпалюють потенціали дії; вони реагують на зміни світла градуйованими рецепторними потенціалами (деполяризація або гіперполяризація). Незважаючи на це, фоторецептори все ще виділяють глутамат на біполярні клітини. Кількість вивільненого глутамату змінюється разом з мембранним потенціалом, тому гіперполяризація призведе до зменшення вивільнення глутамату. Фоторецептори гіперполяризуються на світлі і деполяризуються в темряві. На графіках, використовуваних в цьому уроці, потенціал пускової мембрани буде залежати від початкового стану освітлення.
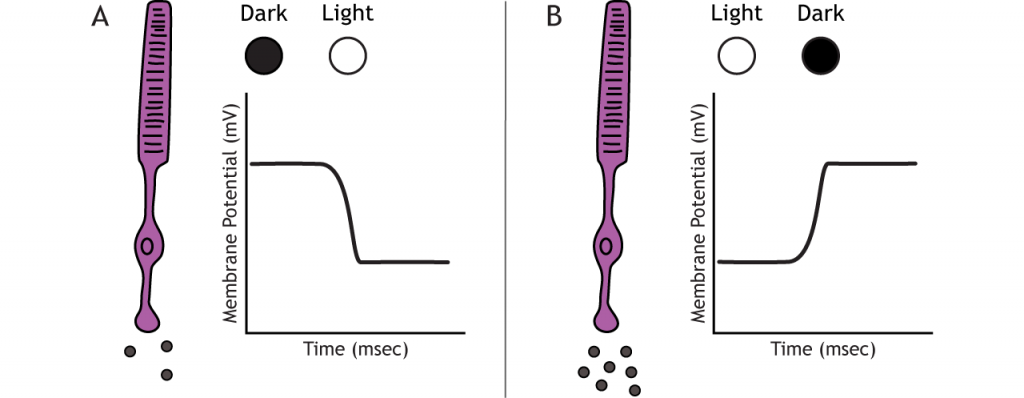
У темряві фоторецептор має мембранний потенціал, який більш деполяризований, ніж «типовий» нейрон, який ми розглядали в попередніх розділах; мембранний потенціал фоторецепторів становить приблизно -40 мВ. Фоторецептори мають відкриті катіонні канали, що дозволяють приплив натрію і кальцію в темний час доби. Ці канали закриті наявністю циклічного GMP (cGMP), молекули важливої в каскадах другого месенджера, яка присутня в фоторецепторі в темряві.
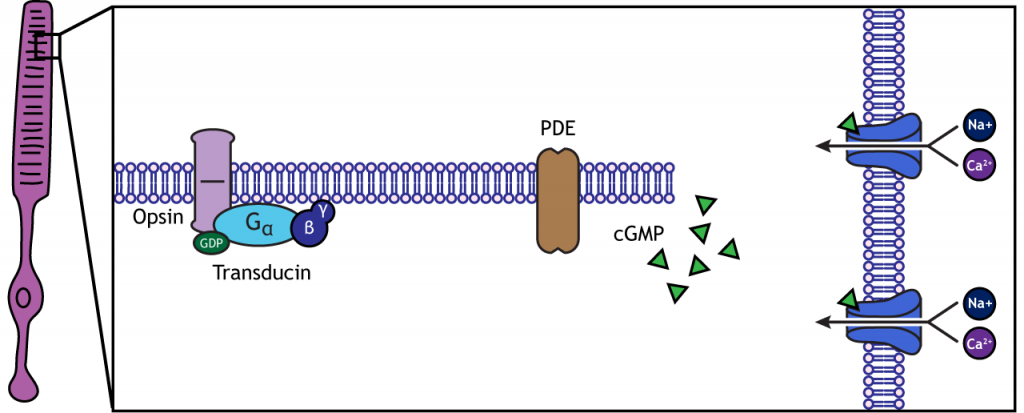
Коли фоторецептор рухається в світло, клітина гіперполяризується. Світло потрапляє в око, досягає фоторецепторів і викликає конформаційні зміни в спеціальному білку, який називається опсіном. Ця зміна активує G-білок, який називається трансдукін, який потім активує білок, який називається фосфодіестеразою (PDE). PDE розбиває cGMP до GMP, а іонні канали CGMP, які були відкриті в темряві, закриваються. Зменшення потоку катіонів в клітину змушує фоторецептор гіперполяризуватися.
Анімація 19.1. Світло, що досягає фоторецептора, викликає конформаційну зміну білка опсину, який активує перетворення G-білка. Предукін активує фосфодіестеразу (PDE), яка перетворює cGMP в GMP. Без цГМП катіонні канали закриваються, зупиняючи приплив позитивних іонів. Це призводить до гіперполяризації клітини. «Фотоперетворення» Кейсі Хенлі ліцензовано на умовах Ліцензії Creative Commons Із Зазначенням Авторства Некомерційна Частка На Тих Самих Умовах (CC BY-NC-SA) 4.0 Міжнародна. Перегляд статичного зображення анімації.
Передача інформації в межах Retina
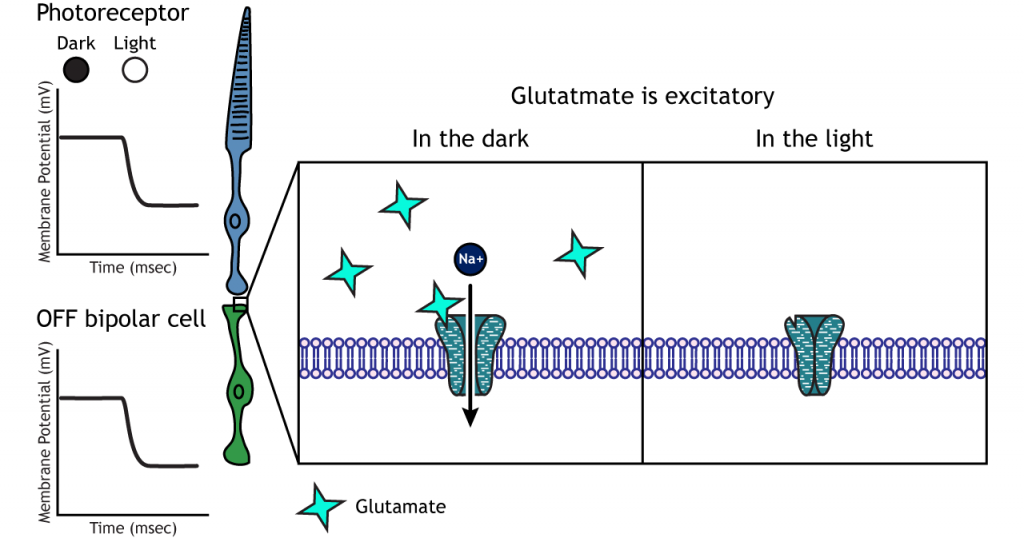
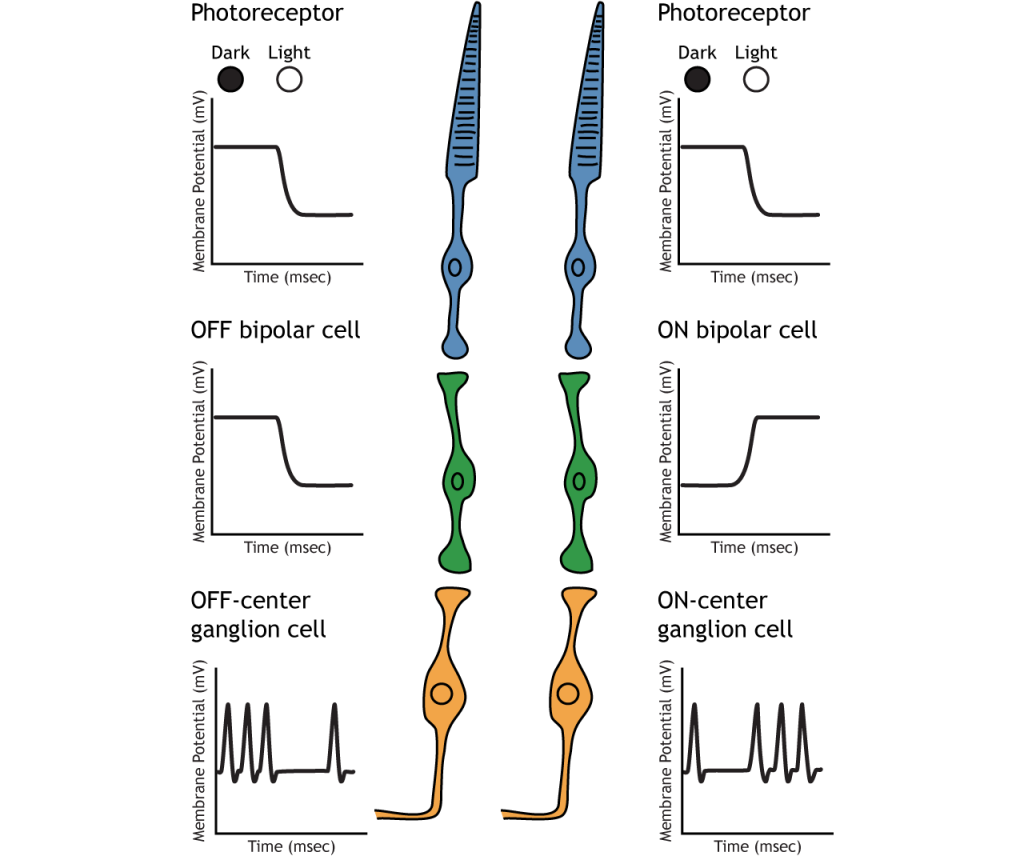
Фоторецептори синапсують на біполярні клітини сітківки. Існує два типи біполярних клітин: OFF і ON. Ці клітини реагують протилежними способами на глутамат, що виділяється фоторецепторами, оскільки вони експресують різні глутаматні рецептори. Як і фоторецептори, біполярні клітини не мають потенціалу вогневої дії і реагують лише градуйованими постсинаптичними потенціалами.
OFF біполярні клітини
У OFF біполярних клітинок глутамат, що виділяється фоторецептором, є збудливим. OFF біполярні клітини експресують іонотропні глутамат рецептори У темряві глутамат, що виділяється фоторецептором, активує іонотропні рецептори, а натрій може надходити в клітину, деполяризуючи мембранний потенціал. На світлі відсутність глутамату змушує іонотропні рецептори закриватися, запобігаючи приплив натрію, гіперполяризуючи мембранний потенціал.
ON Біполярні клітини
У біполярних клітині ON глутамат, що виділяється фоторецептором, є інгібуючим. НА біполярні клітини експресують метаботропні глутаматні рецептори. У темряві глутамат, що виділяється фоторецептором, активує метаботропні рецептори, а G-білки закривають катіонні канали в мембрані, зупиняючи приплив натрію і кальцію, гіперполяризуючи мембранний потенціал. У світлі відсутність глутамату призводить до того, що іонні канали відкриті і дозволяють приплив катіонів, деполяризуючи мембранний потенціал.
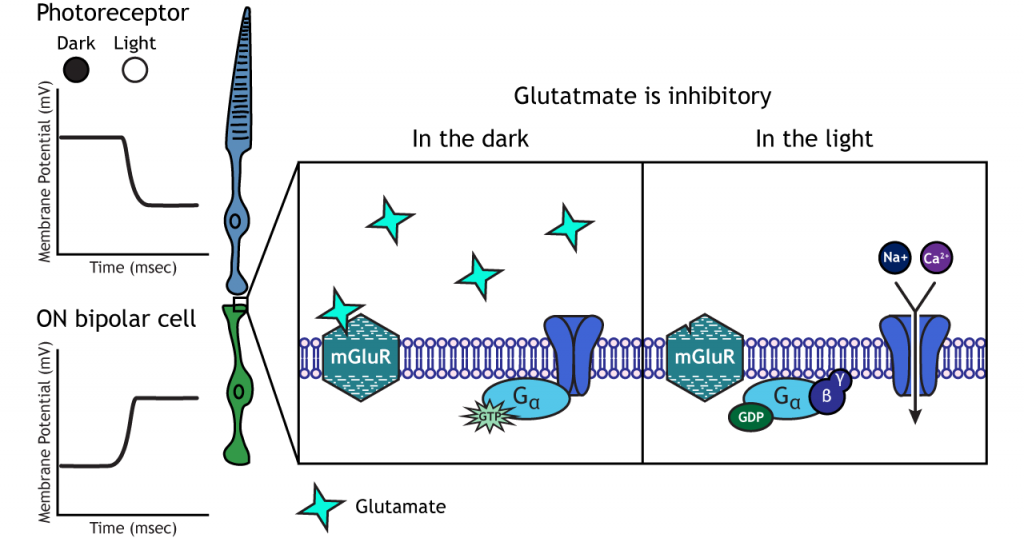
гангліозні клітини
OFF та ON біполярні клітини синапсу на позацентрових та он-центральних гангліозних клітинам відповідно. Гангліозні клітини є єдиним типом клітин для надсилання інформації з сітківки, і вони також є єдиною клітиною, яка запускає потенціали дії. Клітини гангліїв стріляють при будь-яких умовах освітлення, але саме відносна швидкість стрільби кодує інформацію про світло. Перехід від темного до світлого призведе до того, що поза центром гангліозні клітини зменшать швидкість стрільби, а в центрі гангліозних клітин збільшити швидкість стрільби.
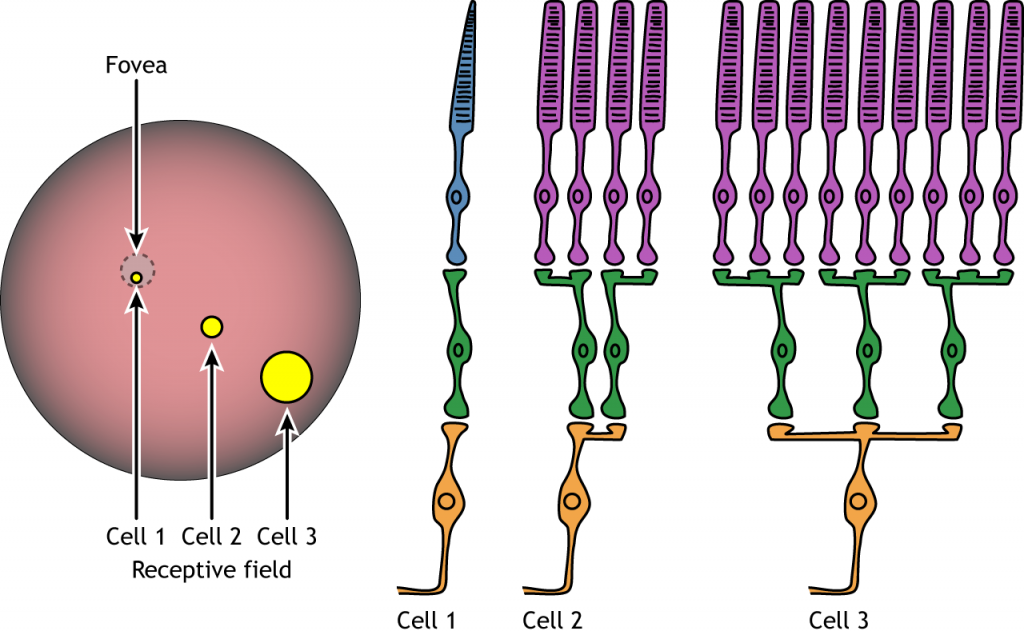
Сприйнятливі поля
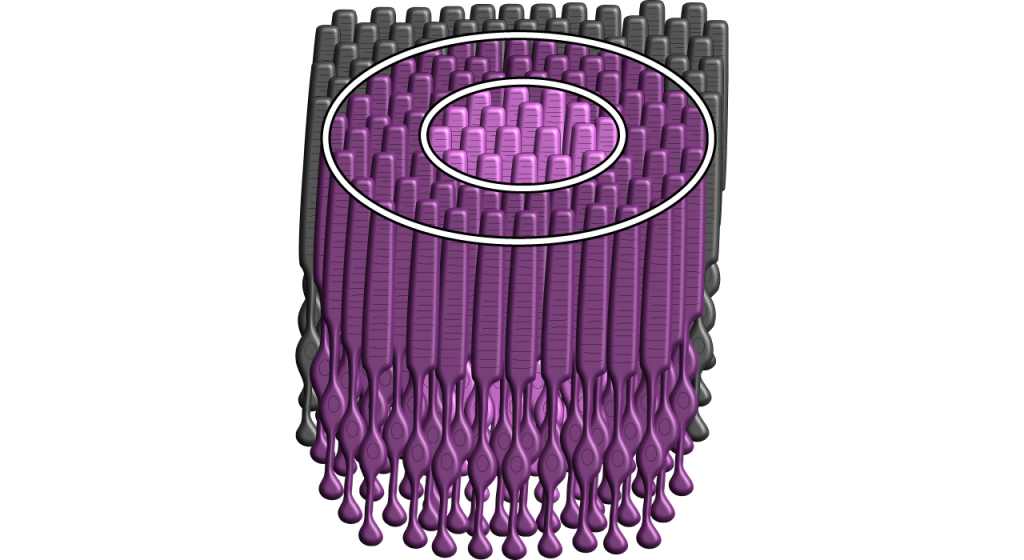
Кожна біполярна і гангліозна клітина реагує на світловий подразник в певній області сітківки. Ця область сітківки є сприйнятливим полем клітини. Сприйнятливі поля в сітківці мають круговий характер.
Розмір сприйнятливого поля може варіюватися. Ямка має менші сприйнятливі поля, ніж периферична сітківка. Розмір залежить від кількості фоторецепторів, які синапсують на даній біполярній клітині, і кількості біполярних клітин, які синапсують на даній гангліозної клітині, також називається величиною конвергенції.
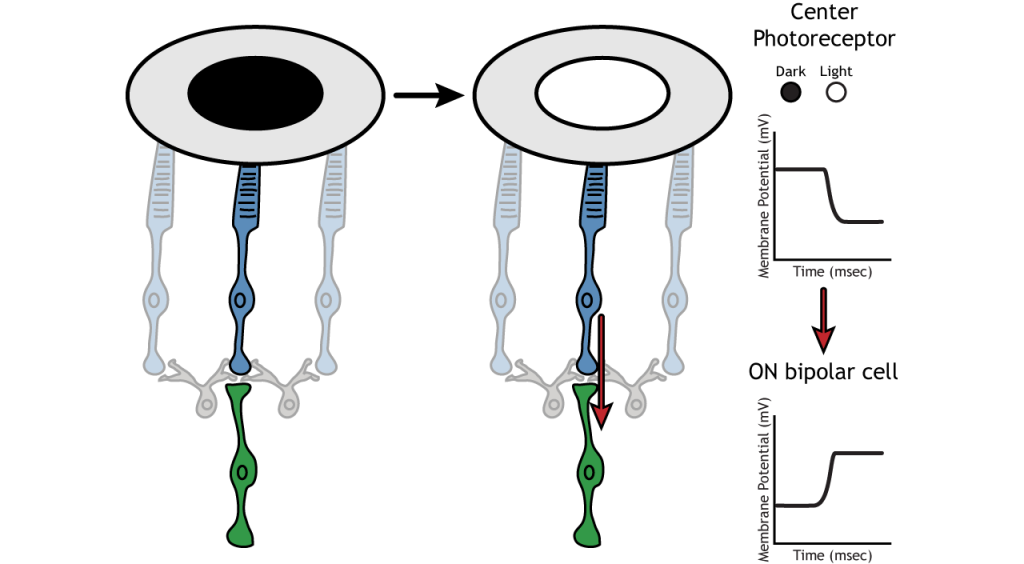
Приклад сприйнятливого поля
Давайте використаємо приклад біполярної клітини ON, щоб подивитися на структуру сприйнятливих полів сітківки. Біполярні і гангліозні сприйнятливі поля поділяються на дві області: центр і об'ємний. Центр рецептивного поля є результатом прямої іннервації між фоторецепторами, біполярними клітинами і гангліозними клітинами. Якщо світлова пляма охоплює центр рецептивного поля, біполярна клітина ON деполяризується, як обговорювалося вище; світло потрапляє на фоторецептор, він гіперполяризується, зменшуючи вивільнення глутамату. Менше глутамату призводить до меншого пригнічення ON біполярної клітини, і вона деполяризується.
Об'ємна частина рецептивного поля є результатом непрямої комунікації між нейронами сітківки через горизонтальні та амакринові клітини. Об'ємне оточення також має протилежну дію на біполярну або гангліозну клітину порівняно з ефектом центральної області. Якщо світлова пляма охоплює об'ємну частину, біполярна клітина ON відповість гіперполяризацією. Світло призведе до гіперполяризації фоторецептора в об'ємі. Це призведе до того, що горизонтальна клітина також гіперполяризується. Горизонтальні клітини мають інгібуючі синаптичні ефекти, тому гіперполяризація в горизонтальній клітині призведе до деполяризації в центрі фоторецептора. Потім центральний фоторецептор спричинить гіперполяризацію в біполярній клітині ON. Ці ефекти імітують ті, що спостерігаються, коли центр знаходиться в темряві. Тож незважаючи на те, що центральний фоторецептор безпосередньо не відчуває зміни умов освітлення, нейрони реагують так, ніби вони рухаються до темряви.
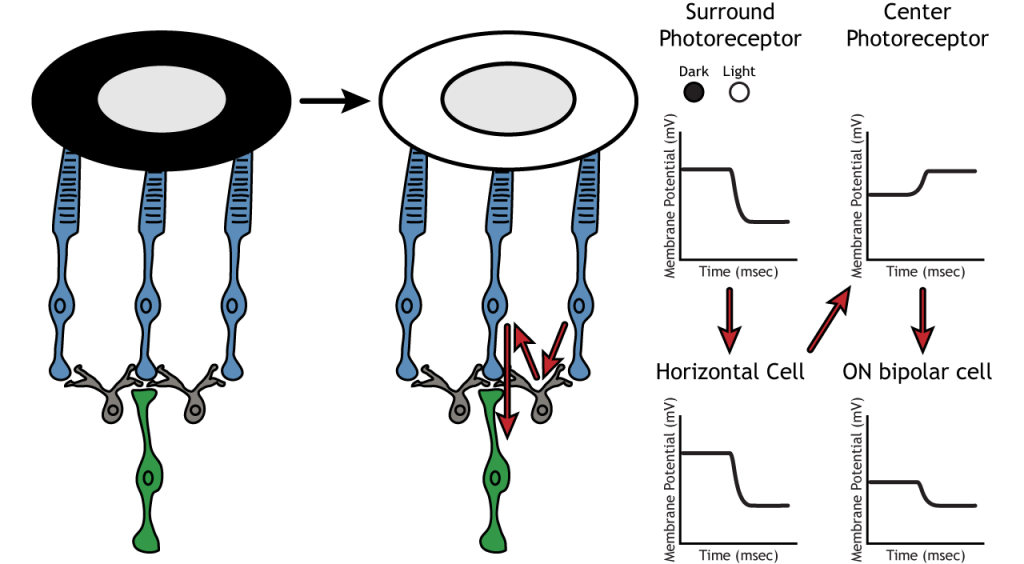
Бічне гальмування
Центральна об'ємна структура рецептивного поля має вирішальне значення для бокового гальмування. Бічне гальмування - це здатність сенсорних систем посилювати сприйняття країв подразників. Важливо відзначити, що фоторецептори, які знаходяться в оточенні однієї біполярної клітини, також будуть знаходитися в центрі іншої біполярної клітини. Це призводить до прямого синаптичного впливу на одну біполярну клітину, одночасно надаючи непрямий вплив на іншу біполярну клітину.
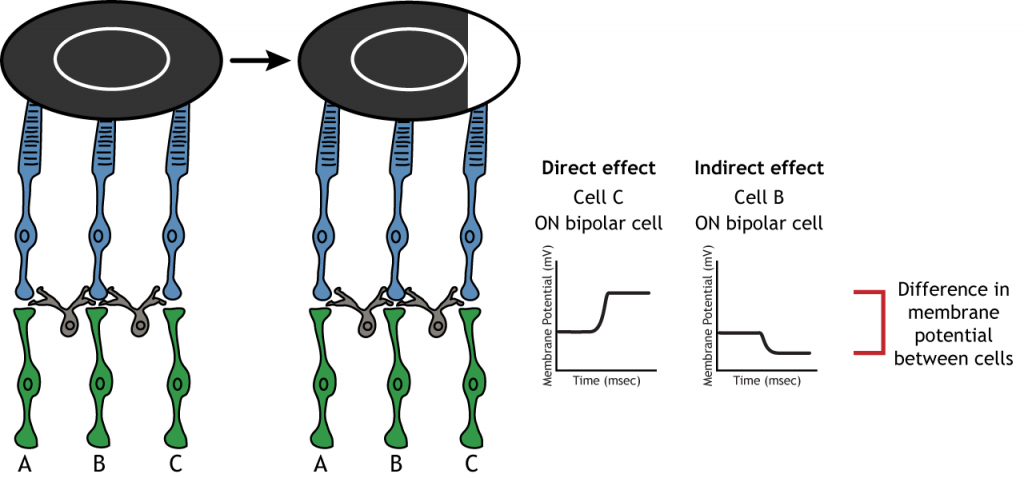
Хоча деякі зображення, використовувані тут, спростять сприйнятливе поле до однієї клітини в центрі і пари в об'ємі, важливо пам'ятати, що фоторецептори покривають всю поверхню сітківки, а сприйнятливе поле двовимірне. Залежно від рівня конвергенції на біполярних і гангліозних клітині рецептивні поля можуть містити безліч фоторецепторів.
Ключові виноси
- Фоторецептори і біполярні клітини не розпалюють потенціали дії
- Фоторецептори гіперполяризуються на світлі
- НА біполярні клітини експресують інгібуючі метаботропні глутаматні рецептори
- OFF біполярні клітини експресують збудливі іонотропні глутамат рецептори
- Сприйнятливі поля круглі, мають центр і об'єм, і відрізняються за розміром
- Структура сприйнятливого поля дозволяє відбуватися бічне гальмування
Перевірте себе!
Інтерактивний елемент H5P був виключений з цієї версії тексту. Ви можете переглянути його онлайн тут:
https://openbooks.lib.msu.edu/neuroscience/?p=438#h5p-18
Додатковий відгук
- Порівняйте і контрастні стрижні і шишки.
- Порівняйте і порівняйте ямку і диск зорового нерва.
Відповіді
Відео-версія уроку

Елемент YouTube був виключений з цієї версії тексту. Ви можете переглянути його онлайн тут: https://openbooks.lib.msu.edu/neuroscience/?p=438