10.4: Будова та функції клітинних геномів
- Page ID
- 3903

Цілі навчання
- Визначити ген і генотип і диференціювати генотип від фенотипу
- Опишіть структуру хромосом і упаковку
- Порівняйте прокаріотичні та еукаріотичні хромосоми
- Поясніть, чому екстрахромосомна ДНК важлива в клітині
До цих пір ми обговорювали будову та функцію окремих шматочків ДНК та РНК. У цьому розділі ми обговоримо, як весь генетичний матеріал організму, який спільно називають його геном, організований всередині клітини. Оскільки генетика організму в значній мірі диктує його особливості, не повинно дивуватися, що організми відрізняються розташуванням своєї ДНК і РНК.
Генотип проти фенотипу
Вся клітинна діяльність кодується в ДНК клітини. Послідовність основ всередині молекули ДНК представляє генетичну інформацію клітини. Сегменти молекул ДНК називаються генами, а окремі гени містять інструкційний код, необхідний для синтезу різних білків, ферментів або стабільних молекул РНК.
Повна колекція генів, які клітина містить у своєму геномі, називається її генотипом. Однак клітина не експресує всі свої гени одночасно. Замість цього він включає (експресує) або вимикає певні гени, коли це необхідно. Набір генів, що виражаються в будь-який даний момент часу, визначає діяльність клітини та її спостережувані характеристики, іменовані як її фенотип. Гени, які завжди виражені, відомі як складові гени; деякі складові гени відомі як гени господарювання, оскільки вони необхідні для основних функцій клітини.
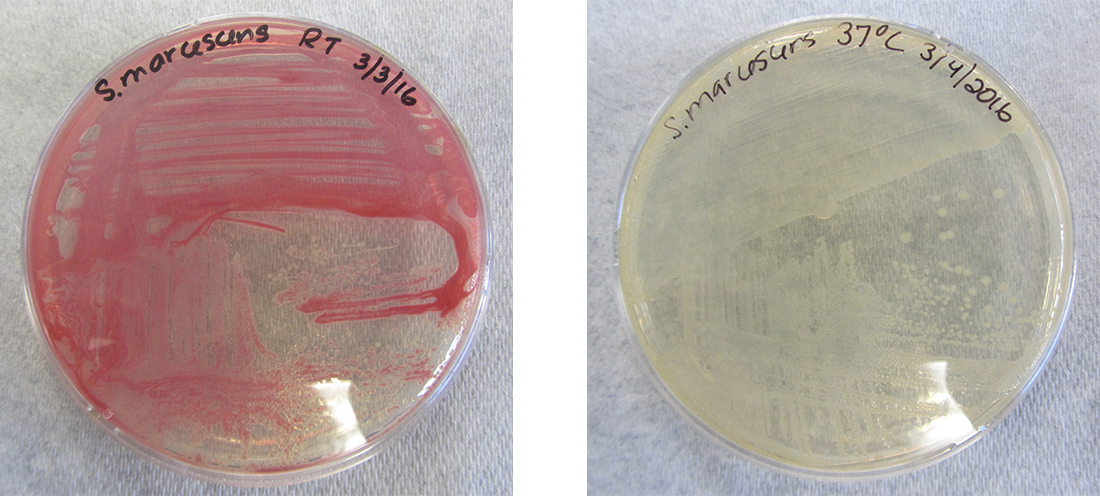
Хоча генотип клітини залишається постійним, фенотип може змінюватися у відповідь на сигнали навколишнього середовища (наприклад, зміни температури або доступності поживних речовин), які впливають на експресію неконституційних генів. Наприклад, оральна бактерія Streptococcus mutans виробляє липкий шар слизу, який дозволяє їй прилипати до зубів, утворюючи зубний наліт; однак гени, які контролюють вироблення шару слизу, виражаються лише в присутності сахарози (столового цукру). Таким чином, поки генотип S. mutans постійний, його фенотип змінюється залежно від наявності та відсутності цукру в його середовищі. Температура також може регулювати експресію генів. Наприклад, грамнегативна бактерія Serratia marcescens, збудник часто асоціюється з лікарняними інфекціями, виробляє червоний пігмент при 28° C, але не при 37° C, нормальній внутрішній температурі людського тіла (рис.\(\PageIndex{1}\)).
Організація генетичного матеріалу
Переважна більшість генома організму організована в хромосоми клітини, які є дискретними структурами ДНК всередині клітин, які контролюють клітинну активність. Нагадаємо, що в той час як еукаріотичні хромосоми розміщені в мембрано-зв'язаному ядрі, більшість прокаріотів містять єдину кругову хромосому, яка знаходиться в області цитоплазми, яка називається нуклеоїдом (див. Унікальні характеристики прокаріотичних клітин). Хромосома може містити кілька тисяч генів.
Організація еукаріотичної хромосоми
Структура хромосом дещо відрізняється між еукаріотичними і прокаріотичними клітинами. Еукаріотичні хромосоми, як правило, лінійні, а еукаріотичні клітини містять кілька різних хромосом. Багато еукаріотичні клітини містять по дві копії кожної хромосоми і, отже, є диплоїдними.
Довжина хромосоми значно перевищує довжину клітини, тому хромосому потрібно упаковувати в дуже маленький простір, щоб поміститися всередині клітини. Наприклад, об'єднана довжина всіх 3 мільярдів пар основ 1 ДНК генома людини вимірювала б приблизно 2 метри, якщо вони повністю розтягнуті, а деякі еукаріотичні геноми у багато разів більші, ніж геном людини. Суперколінг ДНК відноситься до процесу, за допомогою якого ДНК скручується, щоб поміститися всередині клітини. Супернамотування може призвести до того, що ДНК або підмотується (менше одного витка спіралі на 10 пар основ), або перемотується (більше одного обороту на 10 пар основ) з нормального розслабленого стану. Білки, які, як відомо, беруть участь у суперколінгу, включають топоізомерази; ці ферменти допомагають підтримувати структуру перекручених хромосом, запобігаючи перемотування ДНК під час певних клітинних процесів, таких як реплікація ДНК.
Під час упаковки ДНК ДНК-зв'язуючі білки, звані гістонами, виконують різні рівні обгортання ДНК та прикріплення до білків будівельних лісів. Поєднання ДНК з цими прикріпленими білками називають хроматином. У еукаріотів на упаковку ДНК гістонами можуть впливати фактори навколишнього середовища, які впливають на наявність метильних груп на певних цитозинових нуклеотидах ДНК. Вплив факторів навколишнього середовища на упаковку ДНК називається епігенетикою. Епігенетика - ще один механізм регулювання експресії генів без зміни послідовності нуклеотидів. Епігенетичні зміни можуть підтримуватися за допомогою декількох раундів поділу клітин і, отже, можуть бути спадковими.
Перегляньте цю анімацію з навчального центру ДНК, щоб дізнатись більше про упаковку ДНК у еукаріотів.
Організація прокаріотичних хромосом
Хромосоми у бактерій та архей зазвичай кругові, а прокаріотична клітина зазвичай містить лише одну хромосому всередині нуклеоїда. Оскільки хромосома містить лише одну копію кожного гена, прокаріоти гаплоїдні. Як і в еукаріотичних клітині, суперколінг ДНК необхідний для того, щоб геном вписався в прокаріотичну клітину. ДНК в бактеріальної хромосомі розташована в декількох суперспіральних доменах. Як і у випадку з еукаріотами, топоізомерази беруть участь в суперколінгу ДНК. ДНК-гіраза - це тип топоізомерази, що міститься в бактеріях та деяких археях, що допомагає запобігти перемотуванню ДНК. (Деякі антибіотики вбивають бактерії шляхом націлювання на ДНК-гіразу.) Крім того, гістоноподібні білки зв'язують ДНК і допомагають в упаковці ДНК. Інші білки зв'язуються з походженням реплікації, місцем розташування в хромосомі, де ініціюється реплікація ДНК. Оскільки різні ділянки ДНК упаковані по-різному, деякі ділянки хромосомної ДНК більш доступні для ферментів і, таким чином, можуть бути використані більш легко як шаблони для експресії генів. Цікаво, що було показано, що кілька бактерій, включаючи Helicobacter pylori та Shigella flexneri, викликають епігенетичні зміни у своїх господарів після зараження, що призводить до ремоделювання хроматину, що може спричинити довгостроковий вплив на імунітет господаря. 2
Вправа\(\PageIndex{1}\)
- У чому різниця між генотипом клітини та її фенотипом?
- Як ДНК поміщається всередині клітин?
Некодування ДНК
Крім генів, геном також містить багато областей некодуючої ДНК, які не кодують білки або стабільні продукти РНК. Некодуюча ДНК зазвичай зустрічається в областях до початку кодування послідовностей генів, а також в інтергенних областях (тобто послідовності ДНК, розташовані між генами) (рис.\(\PageIndex{2}\)).
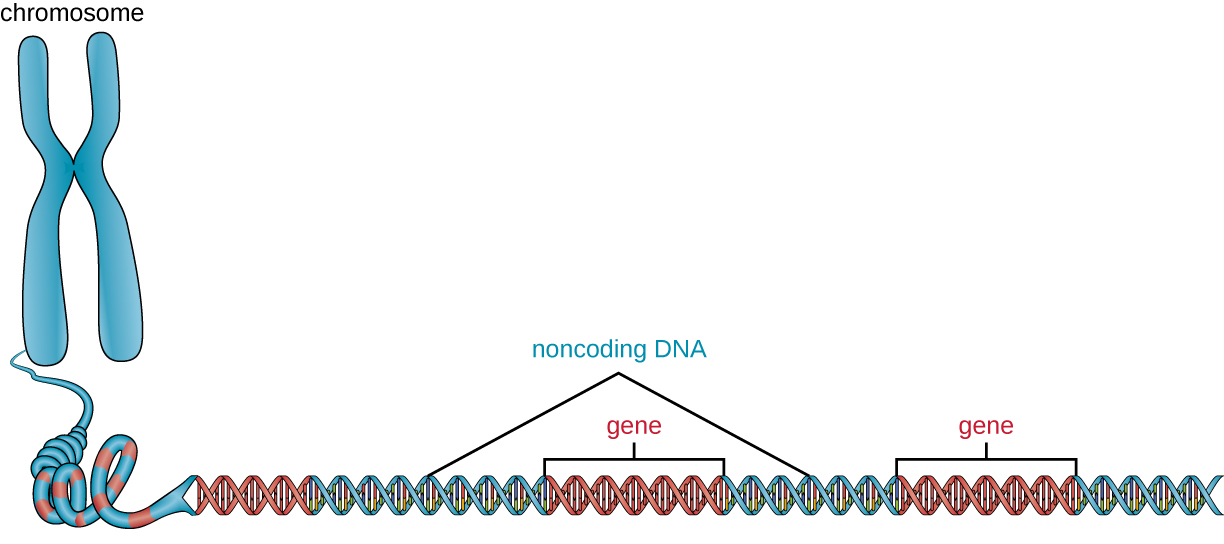
Прокаріоти, як видається, використовують свої геноми дуже ефективно, причому лише в середньому 12% генома захоплюються некодуючими послідовностями. На відміну від цього, некодуюча ДНК може становити близько 98% генома в еукаріотів, як це видно у людини, але відсоток некодуючої ДНК варіюється в різних видах. 3 Ці некодуючі ділянки ДНК колись називали «сміттєвою ДНК»; однак ця термінологія вже не є широко прийнятою, оскільки вчені з тих пір знайшли ролі для деяких з цих регіонів, багато з яких сприяють регулюванню транскрипції або перекладу шляхом виробництва малих некодуючі молекули РНК, упаковка ДНК та хромосомна стабільність. Хоча вчені можуть не до кінця зрозуміти ролі всіх некодуючих областей ДНК, прийнято вважати, що вони мають цілі всередині клітини.
Вправа\(\PageIndex{2}\)
Яка роль некодує ДНК?
Екстрахромосомні ДНК
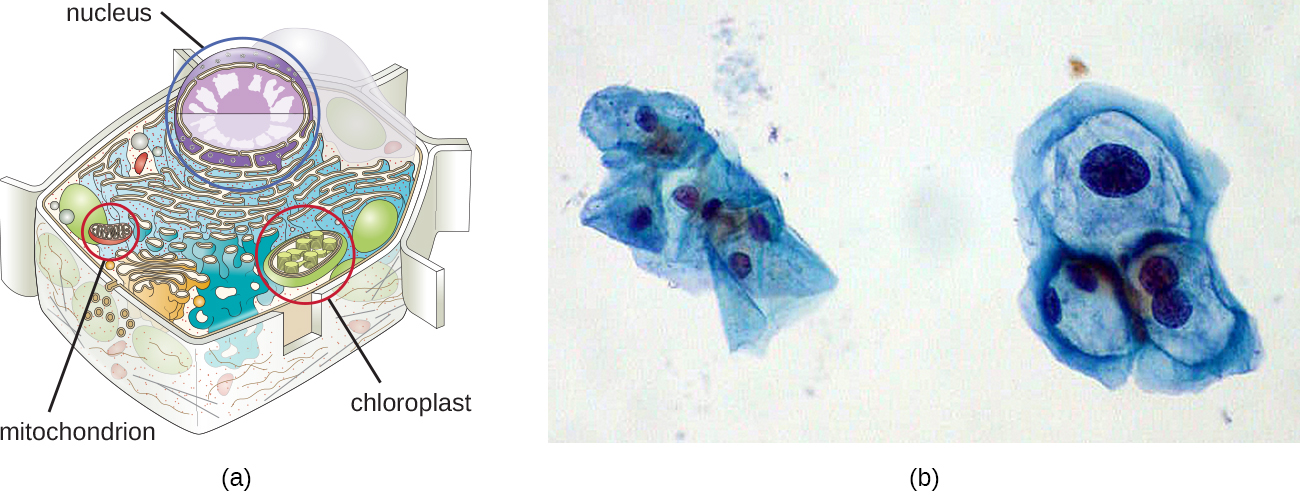
Хоча більшість ДНК міститься в хромосомах клітини, багато клітин мають додаткові молекули ДНК поза хромосом, які називаються екстрахромосомною ДНК, які також є частиною її генома. Геноми еукаріотичних клітин також включали б хромосоми з будь-яких органел, таких як мітохондрії та/або хлоропласти, які ці клітини підтримують (рис.\(\PageIndex{3}\)). Зміст кругових хромосом у цих органелах є пережитком їх прокаріотичного походження та підтримує ендосимбіотичну теорію (див. Основи сучасної теорії клітин). У деяких випадках геноми певних ДНК-вірусів також можуть підтримуватися незалежно в клітинах-господарях під час латентної вірусної інфекції. У цих випадках ці віруси є ще однією формою екстрахромосомної ДНК. Наприклад, вірус папіломи людини (ВПЛ) може підтримуватися в інфікованих клітині таким чином.
Крім хромосом, деякі прокаріоти також мають менші петлі ДНК, які називаються плазмідами, які можуть містити один або кілька генів, не необхідних для нормального росту (рис. 3.3.1). Бактерії можуть обмінюватися цими плазмідами з іншими бактеріями в процесі, відомому як горизонтальний перенос генів (HGT). Обмін генетичного матеріалу на плазміди іноді забезпечує мікроби новими генами, корисними для росту і виживання в особливих умовах. У деяких випадках гени, отримані з плазмід, можуть мати клінічні наслідки, кодуючи фактори вірулентності, які дають мікробу здатність викликати захворювання або робити мікроб стійким до певних антибіотиків. Плазміди також широко використовуються в генній інженерії та біотехнології як спосіб переміщення генів з однієї клітини в іншу. Роль плазмід у горизонтальному переносі генів та біотехнології буде розглянуто далі в Механізмах мікробної генетики та сучасних застосуваннях мікробної генетики.
Вправа\(\PageIndex{3}\)
Як плазміди беруть участь у резистентності до антибіотиків?
Смертельні плазміди
Марія, 20-річна студентка антропології з Техасу, нещодавно захворіла в африканській нації Ботсвана, де проводила дослідження в рамках програми навчання за кордоном. Дослідження Марії були зосереджені на традиційних африканських методах дублення шкур для виробництва шкіри. Протягом трьох тижнів вона відвідувала шкіряний завод щодня протягом декількох годин, щоб спостерігати і брати участь в процесі засмаги. Одного разу після повернення з шкіряного заводу у Марії розвинулася лихоманка, озноб і головний біль, а також біль у грудях, болі в м'язах, нудота та інші грипоподібні симптоми. Спочатку вона не була стурбована, але коли її лихоманка зросла і вона почала кашляти кров, її африканська приймаюча сім'я занепокоїлася і кинула її до лікарні, де її стан продовжував погіршуватися.
Дізнавшись про її недавню роботу на шкіряному заводі, лікар запідозрив, що Марія піддалася сибірській виразці. Він замовив рентген грудної клітки, зразок крові та спинний кран, і негайно почав її на курс внутрішньовенного пеніциліну. На жаль, лабораторні аналізи підтвердили передбачуваний діагноз лікаря. Рентген грудної клітки Марії показав плевральний випіт, скупчення рідини в просторі між плевральними оболонками, а грамова пляма її крові виявила наявність грампозитивних, стрижнеподібних бактерій в коротких ланцюгах, узгоджених з Bacillus anthracis. Також було показано, що кров та бактерії присутні в її спинномозковій рідині, що свідчить про те, що інфекція прогресувала до менінгіту. Незважаючи на підтримуючу терапію і агресивну антибіотикотерапію, Марія впала в стан, що не реагує, і через три дні померла.
Сибірська виразка - захворювання, викликане введенням в організм ендоспор з грампозитивної бактерії B. anthracis. Після зараження у пацієнтів зазвичай розвивається менінгіт, часто з летальними наслідками. У випадку Марії вона вдихала ендоспори, обробляючи шкури тварин, які були заражені.
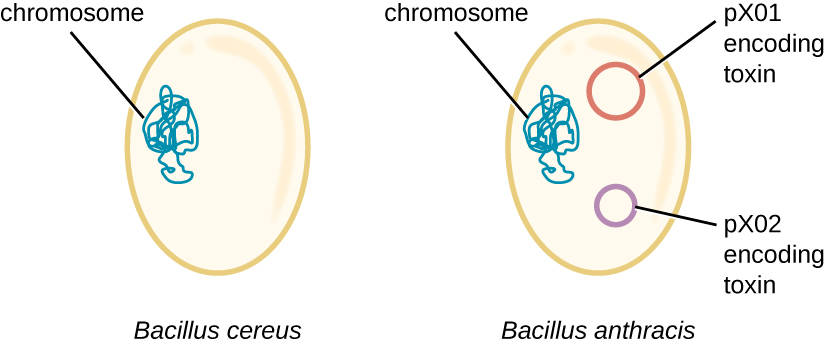
Геном B. anthracis ілюструє, як невеликі структурні відмінності можуть призвести до великих відмінностей у вірулентності. У 2003 році були секвеновані і порівняли геноми B. anthracis і Bacillus cereus, аналогічної, але менш патогенної бактерії того ж роду. 4 Дослідники виявили, що послідовності генів 16S рРНК цих бактерій більш ніж на 99% ідентичні, що означає, що вони насправді є членами одного виду, незважаючи на їх традиційну класифікацію як окремі види. Хоча їх хромосомні послідовності також виявили велику схожість, було виявлено, що кілька факторів вірулентності B. anthracis кодуються на двох великих плазмідах, не знайдених у B. cereus. Плазміда pX01 кодує трикомпонентний токсин, який пригнічує імунну систему господаря, тоді як плазміда Px02 кодує капсульний полісахарид, який додатково захищає бактерію від імунної системи господаря (рис.\(\PageIndex{4}\)). Оскільки B. cereus не вистачає цих плазмід, він не виробляє цих факторів вірулентності, і хоча він все ще є патогенним, він, як правило, пов'язаний з легкими випадками діареї, від якої організм може швидко відновитися. На жаль для Марії, наявність цих плазмід, що кодують токсин, в B. anthracis надає їй смертельну вірулентність.
Вправа\(\PageIndex{4}\)
Як ви думаєте, що трапиться з патогенністю B. anthracis, якщо він втратив одну або обидві свої плазміди?
Клінічна спрямованість: Дозвіл
Протягом 24 годин результати діагностичного тест-аналізу зразка калу Алекса виявили, що він позитивний для теплолабільного ентеротоксину (ЛТ), теплостійкого ентеротоксину (ST) та фактора колонізації (CF), що підтверджує підозру лікаря-лікарні на ЕТЕК. Під час спостереження з сімейним лікарем Алекса цей лікар зазначив, що симптоми Алекса не швидко вирішуються, і він відчував дискомфорт, який заважав йому повернутися до занять. Сімейний лікар призначив Алексу курс ципрофлоксацину для усунення його симптомів. На щастя, ципрофлоксацин усунув симптоми Алекса протягом декількох днів.
Алекс, ймовірно, отримав свою інфекцію від вживання забрудненої їжі або води. Індустріально розвинені країни, такі як Мексика, все ще розробляють санітарні практики, які запобігають забрудненню води фекальним матеріалом. Мандрівникам в таких країнах слід уникати вживання недоварених продуктів, особливо м'яса, морепродуктів, овочів і непастеризованих молочних продуктів. Їм також слід уникати використання води, яка не піддавалася лікуванню; це включає питну воду, кубики льоду і навіть воду, що використовується для чищення зубів. Використання бутильованої води для цих цілей - хороша альтернатива. Хороша гігієна (миття рук) також може допомогти запобігти інфекції ETEC. Алекс не був обережним щодо споживання їжі чи води, що призвело до його хвороби.
Симптоми Алекса були дуже схожі на симптоми холери, викликані грамнегативною бактерією Vibrio cholerae, яка також виробляє токсин, подібний до ST і LT. У якийсь момент еволюційної історії ETEC непатогенний штам кишкової палички, подібний до тих, які зазвичай зустрічаються в кишечнику, можливо, придбав гени, що кодують токсини ST і LT від V. cholerae. Той факт, що гени, що кодують ці токсини, кодуються на екстрахромосомних плазмідах в ETEC, підтримує ідею, що ці гени були придбані E. coli і, ймовірно, підтримуються в бактеріальних популяціях шляхом горизонтального перенесення генів.
вірусні геноми
Вірусні геноми проявляють значну різноманітність у структурі. Деякі віруси мають геноми, які складаються з ДНК як їх генетичного матеріалу. Ця ДНК може бути одноцепочечной, на прикладі парвовіруси людини, або дволанцюгової, як це видно в герпесвірусах і поксвірусах. Крім того, хоча все клітинне життя використовує ДНК як свій генетичний матеріал, деякі вірусні геноми складаються з одноланцюгових або дволанцюгових молекул РНК, як ми вже обговорювали. Вірусні геноми, як правило, менші за більшість бактеріальних геномів, кодуючи лише кілька генів, оскільки вони покладаються на своїх господарів для виконання багатьох функцій, необхідних для їх реплікації. Різноманітність структур вірусного генома та їх наслідки для життєвих циклів реплікації вірусу більш детально розглядаються в Життєвому циклі вірусу.
Вправа\(\PageIndex{5}\)
Чому вірусні геноми широко різняться серед вірусів?
Розмір геному має значення
Існує велика різниця в розмірах геномів серед різних організмів. Більшість еукаріотів підтримують множинні хромосоми; люди, наприклад, мають 23 пари, даючи їм 46 хромосом. Незважаючи на те, що він великий на 3 мільярди пар основ, геном людини далеко не найбільший геном. Рослини часто підтримують дуже великі геноми, до 150 мільярдів пар основ, і зазвичай є поліплоїдними, маючи кілька копій кожної хромосоми.
Розмір геномів бактерій також значно варіюється, хоча вони, як правило, менші, ніж геноми еукаріотів (рис.\(\PageIndex{5}\)). Деякі бактеріальні геноми можуть бути лише 112000 пар основ. Часто розмір генома бактерії безпосередньо пов'язаний з тим, наскільки бактерія залежить від її господаря для виживання. Коли бактерія покладається на клітину господаря для виконання певних функцій, вона втрачає гени, що кодують здатність виконувати ці функції самостійно. Ці типи бактеріальних ендосимбіонтів нагадують прокаріотичне походження мітохондрій і хлоропластів.
З клінічної точки зору облігатні внутрішньоклітинні патогени також мають невеликі геноми (близько 1 мільйона пар основ). Оскільки клітини господаря забезпечують більшу частину своїх поживних речовин, вони, як правило, мають зменшену кількість генів, що кодують метаболічні функції. Завдяки своїм невеликим розмірам геноми таких організмів, як Mycoplasma genitalium (580 000 пар основ), Chlamydia trachomatis (1,0 мільйона), Rickettsia prowazekii (1,1 мільйона) та Treponema pallidum (1,1 мільйона), були одними з попередніх бактеріальних геномів послідовний. Відповідно, ці збудники викликають уретрит і запалення таза, хламідіоз, висипний тиф, сифіліс.
Тоді як облігатні внутрішньоклітинні збудники мають незвично малі геноми, інші бактерії з великою різноманітністю метаболічних і ферментативних можливостей мають надзвичайно великі бактеріальні геноми. Наприклад, синьогнійна паличка - бактерія, яка зазвичай зустрічається в навколишньому середовищі і здатна рости на широкому спектрі субстратів. Його геном містить 6,3 мільйона пар основ, що дає йому високу метаболічну здатність і здатність виробляти фактори вірулентності, що викликають кілька типів опортуністичних інфекцій.
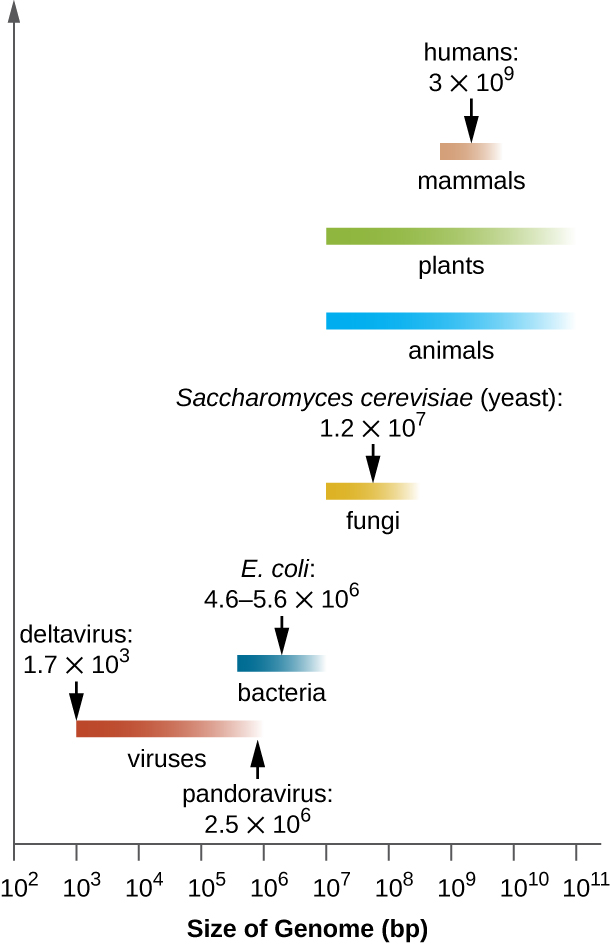
Цікаво, що спостерігалася значна мінливість розміру генома у вірусів, починаючи від 3500 пар основ до 2,5 мільйонів пар основ, що значно перевищує розмір багатьох бактеріальних геномів. Велика варіація, що спостерігається у розмірах вірусного генома, додатково сприяє великій різноманітності вже обговорюваних характеристик вірусного генома.
Відвідайте базу даних геному Національного центру біотехнологічної інформації (NCBI), щоб побачити геноми, які були секвеновані, та їх розміри.
Ключові поняття та резюме
- Весь генетичний вміст клітини - це її геном.
- Гени кодують для білків, або стабільних молекул РНК, кожна з яких виконує певну функцію в клітині.
- Хоча генотип, яким володіє клітина, залишається постійним, експресія генів залежить від умов навколишнього середовища.
- Фенотип - це спостережувані характеристики клітини (або організму) в даний момент часу і є результатом комплементу генів, що використовуються в даний час.
- Більшість генетичного матеріалу організовано в хромосоми, які містять ДНК, яка контролює клітинну діяльність.
- Прокаріоти, як правило, гаплоїдні, зазвичай мають одну кругову хромосому, знайдену в нуклеоїді. Еукаріоти - диплоїдні; ДНК організована в множинні лінійні хромосоми, виявлені в ядрі.
- Суперколінг та упаковка ДНК з використанням білків, що зв'язують ДНК, дозволяє довгим молекулам поміщатися всередині клітини. Еукаріоти і архей використовують гістонові білки, а бактерії використовують різні білки з подібною функцією.
- Прокаріотичні і еукаріотичні геноми обидва містять некодуючу ДНК, функція якої недостатньо вивчена. Деякі некодуючі ДНК, як видається, беруть участь у формуванні малих некодуючих молекул РНК, які впливають на експресію генів; деякі, як видається, відіграють певну роль у підтримці хромосомної структури та в упаковці ДНК.
- Екстрахромосомна ДНК у еукаріотів включає хромосоми, знайдені в органелах прокаріотичного походження (мітохондрії та хлоропласти), які еволюціонували шляхом ендосимбіозу. Деякі віруси також можуть підтримувати себе екстрахромосомно.
- Екстрахромосомна ДНК у прокаріотів зазвичай підтримується як плазміди, які кодують кілька несуттєвих генів, які можуть бути корисними в певних умовах. Плазміди можуть поширюватися через бактеріальне співтовариство шляхом горизонтального перенесення генів.
- Вірусні геноми мають велику варіацію і можуть складатися або з РНК, або ДНК, і можуть бути як подвійними, так і одноланцюжковими.
Виноски
- 1 Національний науково-дослідний інститут геному людини. «Завершення проекту «Геном людини»: Часті запитання». https://www.genome.gov/11006943. Доступно 10 червня 2016 р.
- 2 H. Bierne та ін. «Епігенетика та бактеріальні інфекції». Перспективи Колд-Спрінг-Харбор в медицині 2 № 12 (2012) :a010272.
- 3 Р.Дж. Тафт та ін. «Взаємозв'язок між небілковою ДНК та еукаріотичною складністю». Біоеси 29 № 3 (2007) :288—299.
- Іванова Н.та співавт. «Послідовність геному Bacillus cereus та порівняльний аналіз з Bacillus anthracis». Природа 423 № 6935 (2003) :87—91.