22: Віруси
- Page ID
- 6615
Вірусна класифікація
Оскільки вірусам не вистачає рибосом (і, отже, рРНК), їх не можна класифікувати в рамках схеми класифікації трьох доменів з клітинними організмами. Крім того, доктор Девід Балтімор отримав вірусну схему класифікації, яка фокусується на зв'язку між вірусним геном до того, як він виробляє свою мРНК. Балтіморська схема розпізнає сім класів вірусів.
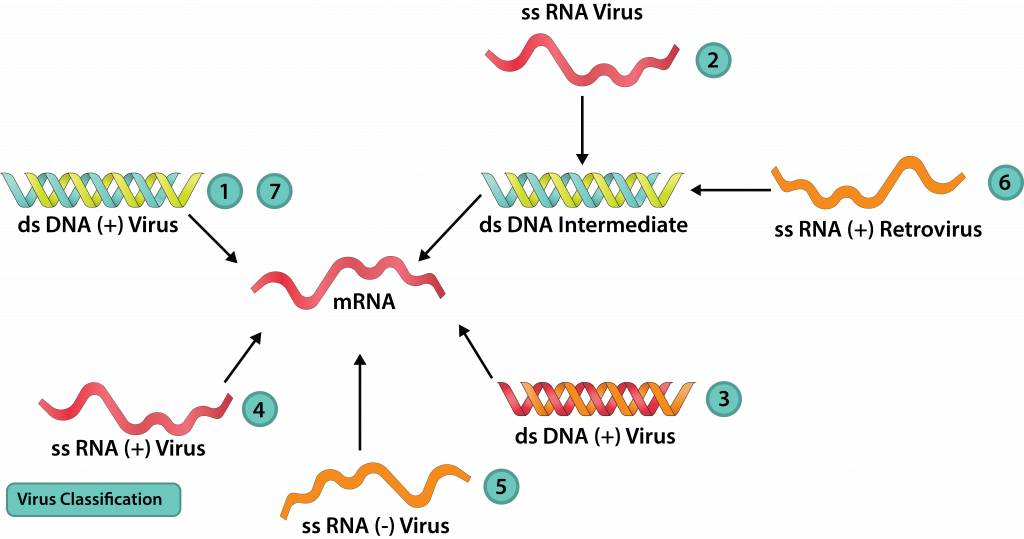
ДНК-віруси
Клас I: дСДНК
ДНК-віруси з геном dsDNA, як бактеріофаги Т4 і лямбда, мають геном точно такий же, як і клітина-господар, якою вони заражають. З цієї причини багато ферментів господаря можуть бути використані для реплікації та/або виробництва білка. Потік інформації слідує звичайним шляхом: dsDNA → мРНК → білок, при цьому ДНК-залежна РНК-полімераза виробляє мРНК і рибосома господаря, що виробляє білок. Реплікація геному, dsDNA → dsDNA, вимагає ДНК-залежної ДНК-полімерази від вірусу або клітини-господаря.
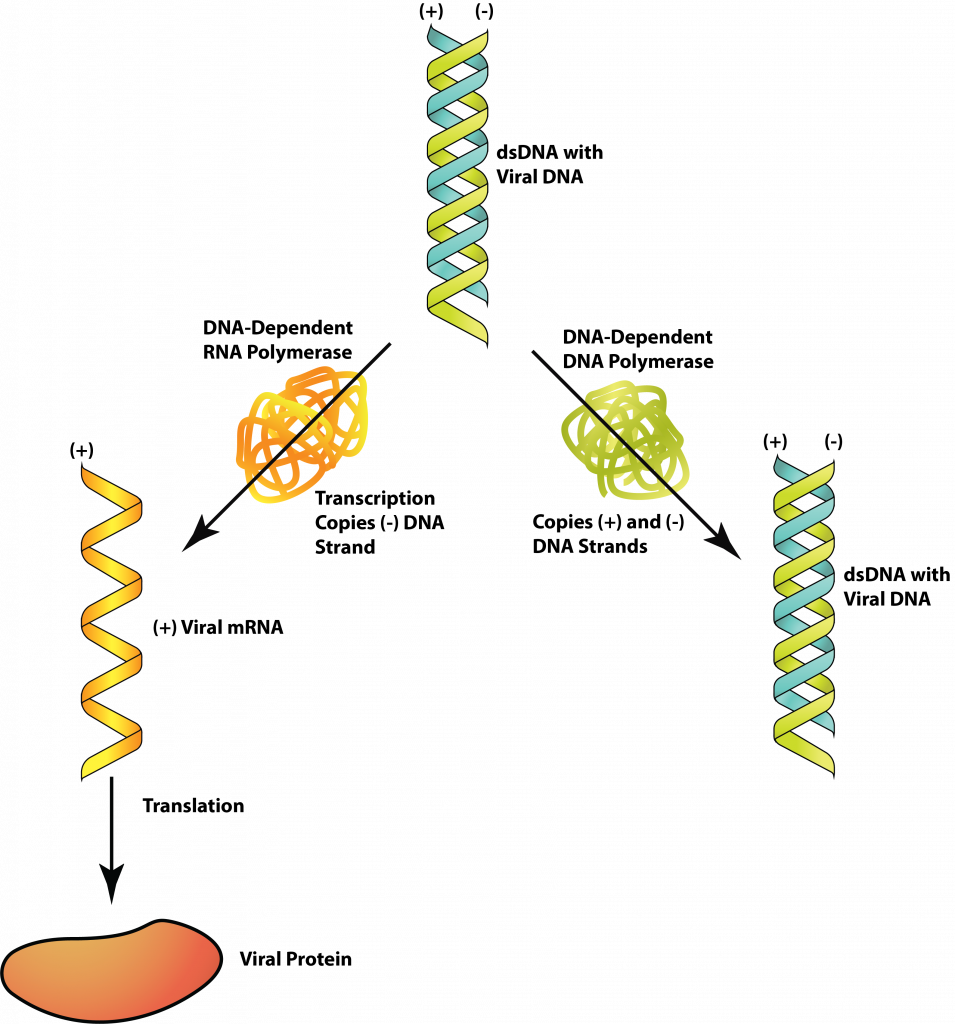
дДНК.
Вірус часто використовує стратегії контролю експресії генів, щоб гарантувати, що конкретні вірусні продукти виробляються в певний час при реплікації вірусу. У випадку Т4 РНК-полімераза господаря зв'язується з вірусною ДНК і починає транскрибувати ранні гени відразу після введення ДНК в клітину. Один з ранніх вірусних білків модифікує РНК-полімеразу господаря так, що він більше не буде розпізнавати промотори господаря взагалі, крім того, щоб перейти до транскрибування генів для вірусних білків середньої стадії. Подальша модифікація (каталізована вірусними білками середньої стадії) додатково модифікувала РНК-полімеразу, щоб вона розпізнавала вірусні гени, які кодують білки пізньої стадії. Це забезпечує впорядковану вироблення вірусних білків.
Реплікація декількох вірусів dsDNA призводить до виробництва конкатемерів, де кілька вірусних геномів пов'язані між собою через короткі одноланцюгові області з термінальними повторами. Оскільки геном упаковується в капсид, вірусна ендонуклеаза розрізає конкатемер до відповідної довжини.
Існує кілька вірусів тварин з геномами dsDNA, таких як віруси віспи та аденовіруси. Герпесвіруси мають кілька помітних особливостей, таких як зв'язок декількох членів з раком і здатність вірусів залишатися в прихованій формі всередині свого господаря. Продуктивна інфекція призводить до вибухонебезпечної вірусної популяції, загибелі клітин і розвитку ознак захворювання, під час яких заражаються нейрони. У нейрони розвивається латентна інфекція, що дозволяє вірусу залишатися непоміченим у господаря. Якщо вірусний геном реактивується, призводить до продуктивної інфекції, що призводить до реплікації вірусу та повторних ознак захворювання.
Клас II: SSDNA
Потік інформації про віруси SSDNA, такі як парвовіруси, все ще буде певною мірою слідувати звичайним шляхом: ДНК → мРНК → білок. Але вірусний геном може або мати ту саму базову послідовність, що і мРНК (плюс-нитка ДНК), або доповнювати мРНК (мінус нитка ДНК). У першому випадку спочатку повинна бути виготовлена ланцюг ДНК, яка доповнює вірусний геном, утворюючи дволанцюгову реплікативну форму (РФ). Це може бути використано як для виробництва вірусних білків, так і як шаблон для копій вірусного генома. Для мінусових ДНК-вірусів геном можна використовувати безпосередньо для отримання мРНК, але ще потрібно буде зробити додаткову копію, щоб служити шаблоном для копій вірусного генома.
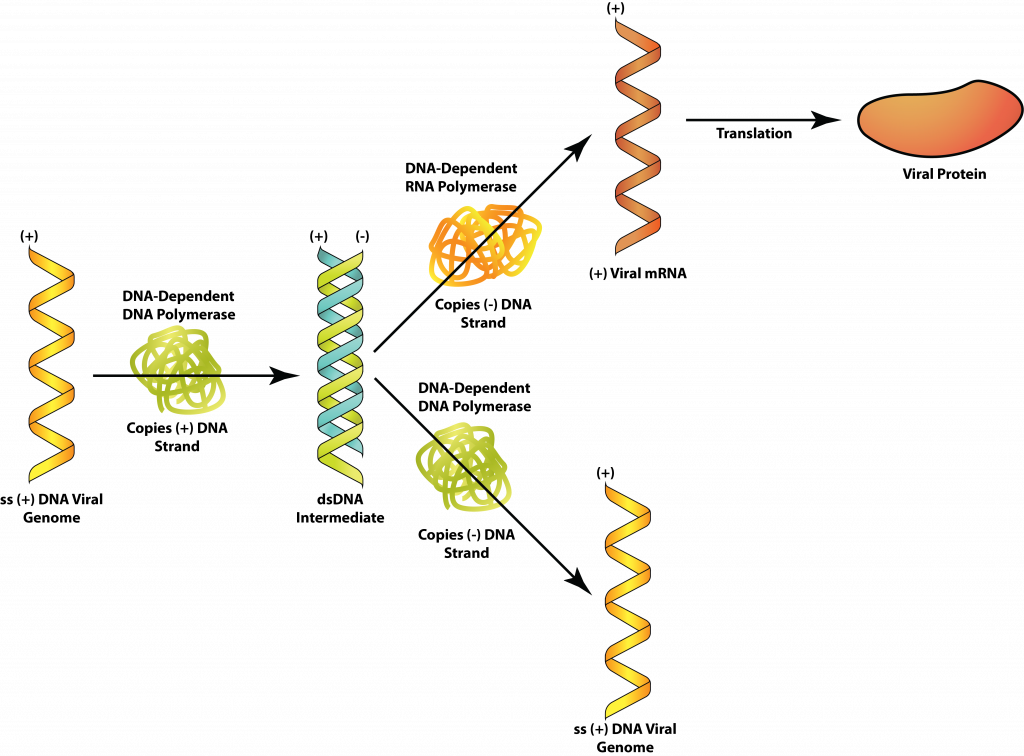
SSDNA.
Реплікативна форма може бути використана для кочення кола реплікації, де одна нитка зарізається і реплікації ферменти використовуються для розширення вільного 3' кінця. Оскільки комплементарна нитка синтезується навколо кругової ДНК, 5-футовий кінець відшаровується, що призводить до зміщеної нитки, яка продовжує рости в довжину.
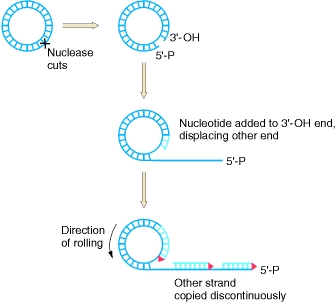
Кочення кола реплікації.
Клас VII: ДНК-віруси, які використовують зворотну транскриптазу
Гепаднавіруси містять геном ДНК, який частково дволанцюговий, але містить одноцепочечную область. Після отримання входу в ядро клітини ферменти клітини-господаря використовуються для заповнення проміжку комплементарними основами для формування замкнутого циклу dsDNA. Генна транскрипція дає плюс-нитку РНК, відому як прегеном, а також вірусний фермент зворотної транскриптази, РНК-залежної ДНК-полімерази. Прегеном використовується як шаблон для зворотної транскриптази для отримання мінус-нитка ДНК геномів, з невеликим шматочком прегенома, який використовується як праймер для отримання дволанцюгової області геномів.
РНК-віруси
Клас III: ДСРНК
Дволанцюгові РНК-віруси заражають бактерії, грибки, рослини та тварин, такі як ротавірус, який викликає діарейні захворювання у людей. Але клітини не використовують dsRNA в жодному зі своїх процесів і мають системи для знищення будь-якої dsRNA, виявленої в клітині. Таким чином, вірусний геном у своїй формі dsRNA повинен бути прихованим або захищеним від клітинних ферментів. Клітинам також не вистачає РНК-залежних РНК-полімераз, необхідних для реплікації вірусного генома, тому вірус повинен забезпечити цей фермент сам. Вірусна РНК-залежна РНК-полімераза діє як транскриптаза для транскрибування мРНК, так і як реплікація для реплікації генома РНК.
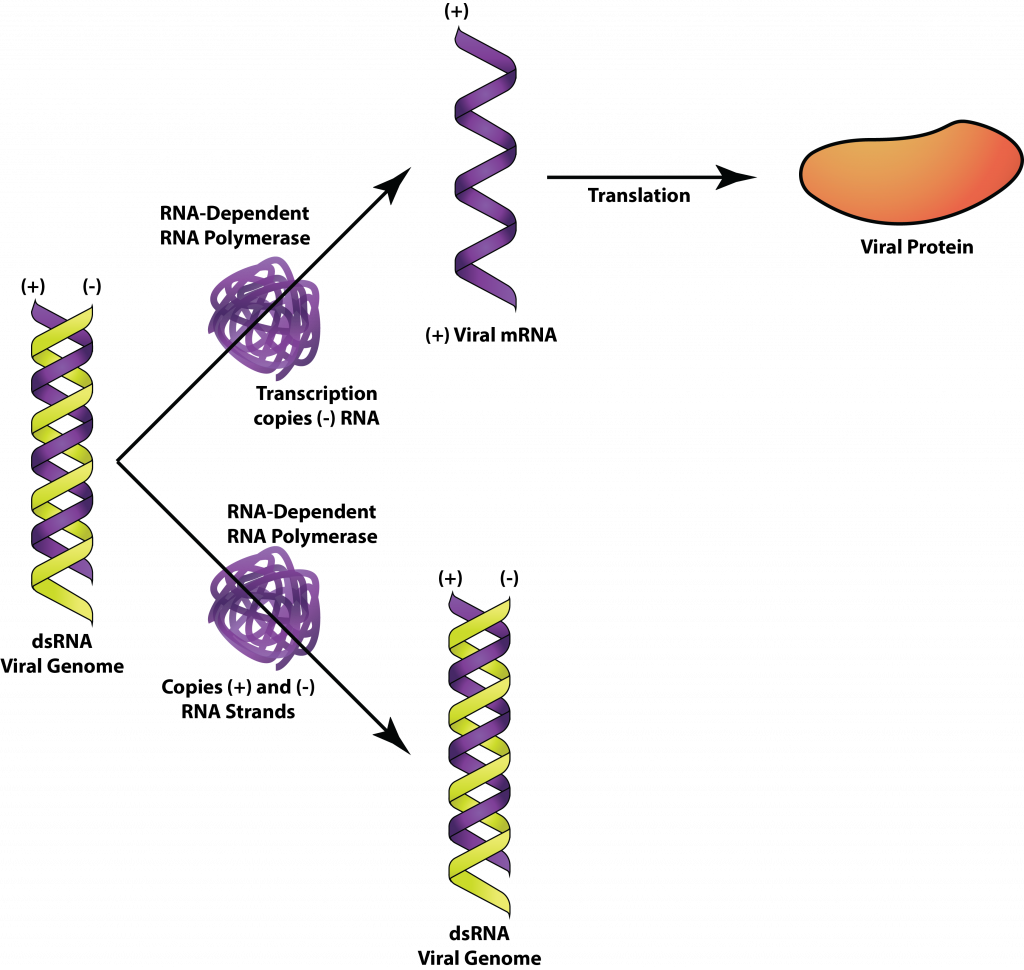
ДСРНК.
Для ротавірусу вірусний нуклеокапсид залишається неушкодженим у цитоплазмі з реплікаційними явищами, що відбуваються всередині, дозволяючи DsRNA залишатися захищеними. Messenger РНК транскрибується з мінус-нитка генома РНК, а потім перекладається рибосомою господаря в цитоплазму. Вірусні білки агрегуються, утворюючи нові нуклеокапсиди навколо реплікази РНК та плюс-нитка РНК. Мінус-ланцюгова РНК потім синтезується репліказою РНК всередині нуклеокапсиду, ще раз забезпечуючи захист генома dsRNA.
Клас IV: +СРНК
Віруси з плюс-ниткою РНК, такі як поліовірус, можуть використовувати свій геном безпосередньо як мРНК з трансляцією рибосомою господаря, що відбувається, як тільки несегментований вірусний геном отримує потрапляння в клітину. Один з експресованих вірусних генів дає РНК-залежну РНК-полімеразу (або РНК-репліказу), яка створює мінус-ниткову РНК з генома плюс-нитка. Мінус-нитка РНК може бути використана як шаблон для більшої кількості плюс-нитки РНК, які можуть бути використані як мРНК або як геноми для знову утворюються вірусів.
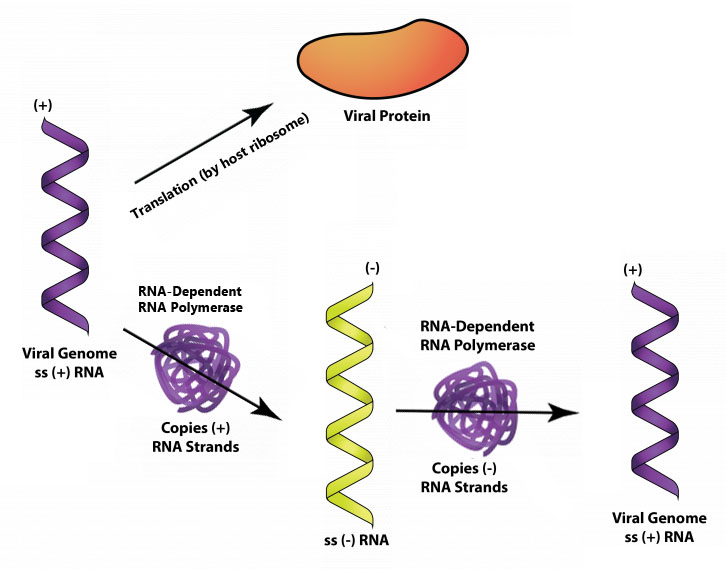
+СРНК.
Переклад генома поліовірусу дає поліпротеїн, великий білок з протеазною активністю, який розщеплюється на три менші білки. Додаткова активність розщеплення в кінцевому підсумку дає всі білки, необхідні для утворення капсиду, а також РНК-залежну РНК-полімеразу.
Формування поліпротеїну, який розрізається на кілька менших білків, ілюструє одну можливу стратегію до проблеми, з якою стикаються багато вірусів+SSRNA - як генерувати кілька білків з несегментованого генома +SSRNA? Інші можливості включають:
- субгеномна мРНК - під час трансляції частини вірусної РНК можуть бути пропущені, в результаті чого білки відрізняються від того, що виробляється з вірусної РНК у повному обсязі.
- рибосомальний зрушення кадрів — рибосома «зчитує» мРНК групами по три нуклеотиди або кодон, які переводяться в одну амінокислоту. Якщо рибосома починається з нуклеотиду #1, тобто один відкритий кадр читання (ORF), в результаті чого утворюється один набір амінокислот. Якби рибосома рухалася вперед, де нуклеотид 2 є початковим нуклеотидом, який був би ORF #2, що призвело до зовсім іншого набору амінокислот. Якби рибосома знову рухатиметься вперед, де нуклеотид 3 є початковим нуклеотидом, який був би ORF #3, що призвело до зовсім іншого набору амінокислот. Деякі віруси мають вірусні гени, які навмисно перекриваються в межах різних ОРФ, що призводить до вироблення різних білків з однієї мРНК.
- механізм читання - вірусний геном може мати стоп-кодони, вбудовані в послідовність. Коли рибосома приходить до зупинки кодон він може або зупинитися, закінчуючи послідовність амінокислот, або він може ігнорувати стоп-кодон, продовжуючи робити довший рядок амінокислот. Для вірусів з механізмом зчитування вони набувають різноманітні білки, маючи стоп-кодони, які періодично ігноруються. Іноді ця функція поєднується зі зміщенням рибосомних кадрів для отримання ще більшої різноманітності вірусних білків.
Клас V: -СРНК
Мінус-ланцюгові РНК-віруси включають багато помітних для людини членів, таких як вірус грипу, вірус сказу та вірус Ебола. Оскільки геном мінусових РНК-вірусів не може використовуватися безпосередньо як мРНК, вірус повинен переносити РНК-залежну РНК-полімеразу всередині свого капсиду. При вході в клітину-господаря плюс-нитки РНК, що генеруються полімеразою, використовуються як мРНК для виробництва білка. Коли потрібні вірусні геноми, плюс-нитка РНК використовуються як шаблони для створення мінус-нитка РНК.
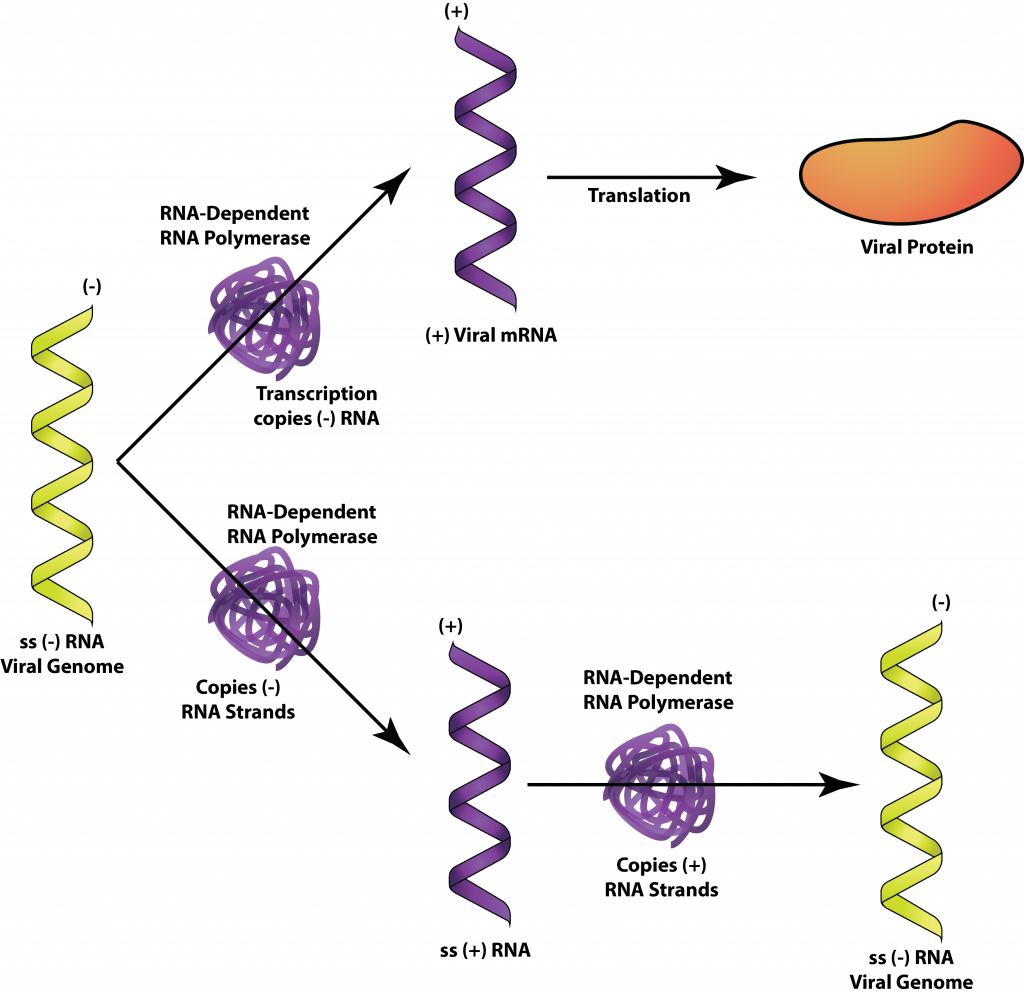
-СРНК.
Клас VI: +SSRNA, ретровіруси
Незважаючи на те, що ретровірусний геном складається з +SSRNA, він не використовується як мРНК. Натомість вірус використовує свою зворотну транскриптазу для синтезу шматочка SSDNA, що доповнює вірусний геном. Зворотна транскриптаза також володіє рибонуклеазной активністю, яка використовується для деградації ланцюга РНК гібрида РНК-ДНК. Нарешті, зворотна транскриптаза використовується як ДНК-полімераза для створення додаткової копії SSDNA, отримуючи молекулу dsDNA. Це дозволяє вірусу вставляти свій геном, у формі dsDNA, в хромосому господаря, утворюючи провірус. На відміну від пророцтва, провірус може залишатися латентним до нескінченності або викликати експресію вірусних генів, що призводить до вироблення нових вірусів. Висічення провірусу не відбувається для експресії генів.
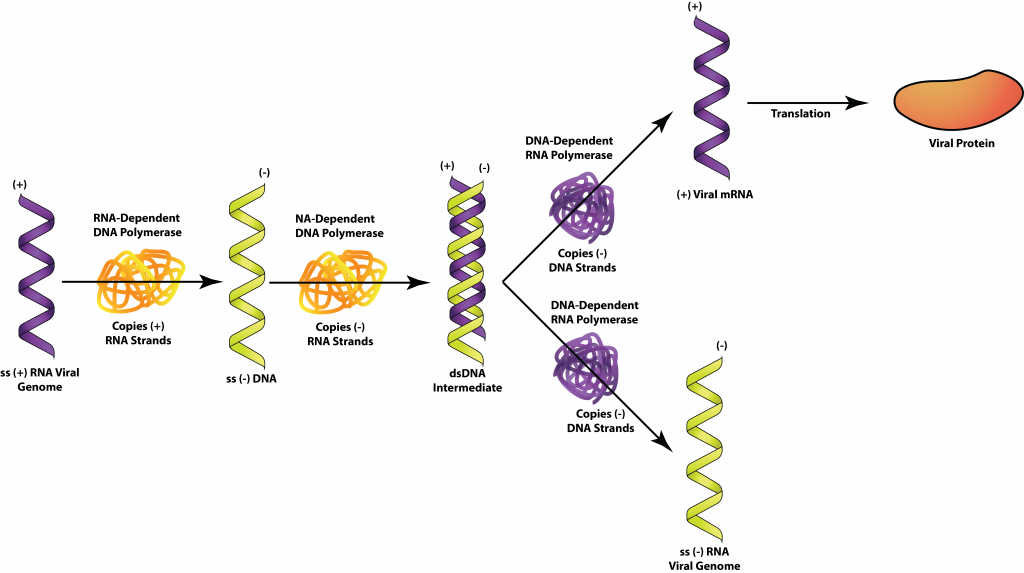
+SSRNA, ретровіруси.
Інші інфекційні агенти
вірусоїди
Віроїди - це невеликі кругові молекули SsRNA, яким не вистачає білка. Ці інфекційні молекули пов'язані з рядом захворювань рослин. Оскільки SsRNA дуже сприйнятлива до ферментативної деградації, вірусоїдна РНК має велику взаємодоповнюючу основу, в результаті чого вірусоїд приймає конфігурацію шпильки, стійку до ферментів. Для реплікації вірусоїди покладаються на рослинну РНК-полімеразу з РНК реплікаційною активністю.
Пріони
Пріони - це інфекційні агенти, яким повністю не вистачає нуклеїнової кислоти будь-якого виду, виготовляючись повністю з білка. Вони пов'язані з безліччю захворювань, в першу чергу у тварин, хоча виявлено пріон, який заражає дріжджі (!). Захворювання включають губчаста енцефалопатія великої рогатої худоби (BSE або «хвороба божевільних коров'ячих»), хвороба Крейцфельда-Якоба у людей та зішкріб у овець.
Білок пріона міститься в нейрони здорових тварин (PRPC або Prion Protein Cellular), з особливою вторинною структурою. Патогенна форма (PrPSc або Prion Protein Scrapie) має різну вторинну структуру і здатна перетворювати PRPC в патогенну форму. Накопичення патогенної форми викликає руйнування мозку і нервової тканини, що призводить до таких симптомів захворювання, як втрата пам'яті, недостатня координація, а згодом і смерть.
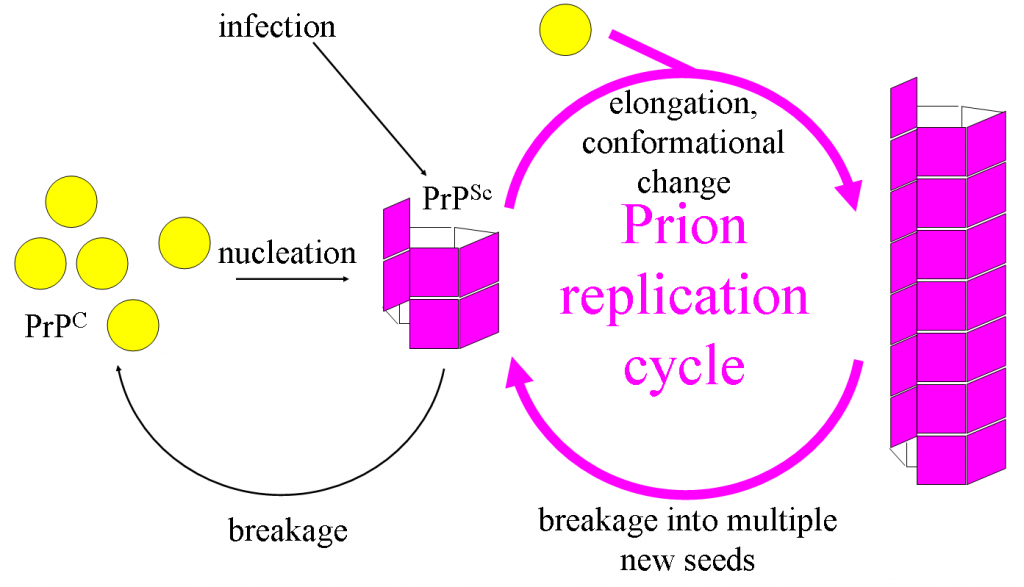
Приони. Джоаннамасел у англійській Вікіпедії [CC BY-SA 3.0], через Wikimedia Commons
Ключові слова
Балтіморська схема, клас I, клас II, клас III, клас IV, клас V, клас VI, клас VII, ДНК-залежна РНК-полімераза, ДНК-залежна ДНК-полімераза, конкатемер, продуктивна інфекція, прихована інфекція, плюс-нитка ДНК/+ ДНК, ДНК-ДНК, DsDNA, реплікаційна форма (РФ), кочення кола реплікації, прегеном, зворотна транскриптаза, РНК-залежна ДНК-полімераза, ДСРНК, РНК-залежна РНК-полімераза, транскриптаза, репліказа, плюс-нитка РНК/+SSRNA, мінус-нитковий РНК/-SSRNA, поліпротеїн, субгеномна мРНК, рибосомальний зрушення кадру, відкритий кадр зчитування (ORF), механізм зчитування, стоп-кодон, ретровірус, рибонуклеаза, провірус, вірусоїдний, пріон, PRPC/Пріон протеїн клітинний, PRPSC/Prion протеїн Scrapie.
Основні питання/цілі
- Що таке Балтіморська система класифікації? Які вірусні характеристики він використовує? Як кожна вірусна група виробляє білки та відтворює їх геном? Звідки беруться необхідні компоненти? (Вірус або клітина господаря) Які модифікації необхідні для вірусів з геном, відмінним від клітини-господаря?
- Яку стратегію використовують віруси dsDNA для контролю експресії генів? Що таке конкатемери? Що таке продуктивні і латентні інфекції?
- Що таке реплікативна форма? Що таке кочення кола реплікації? У чому перевага цих вірусних механізмів?
- Що таке прегеном? Що таке зворотна транскриптаза? Яку роль він відіграє для вірусів класу VII?
- З якими проблемами стикаються віруси DsRNA? Як вони долають ці проблеми? Що таке транскриптаза? Що таке репліказа?
- Як геном використовується вірусами класу IV +SSRNA? Які стратегії використовуються цими вірусами для генерації декількох білків з несегментованого генома?
- Які кроки необхідні для вірусів —SSRNA?
- Чим ретровіруси, як +віруси SSRNA, відрізняються від вірусів IV класу? Що таке рибонуклеаза? Що таке провірус?
- Що таке вірусоїд? Що таке пріон? Як ці кошти викликають захворювання? Як вони відтворюються?
Дослідницькі питання (НЕОБОВ'ЯЗКОВО)
- Чому вчені спочатку були настільки стійкими до ідеї про пріони не вистачає будь-якого типу нуклеїнової кислоти?