8: Вступ до вірусів
- Page ID
- 6621
Віруси, як правило, описуються як облігатні внутрішньоклітинні паразити, ацелюлярні інфекційні агенти, які вимагають наявності клітини-господаря для розмноження. Віруси, які, як було виявлено, заражають всі типи клітин - людей, тварин, рослин, бактерій, дріжджів, археї, найпростіших... деякі вчені навіть стверджують, що вони знайшли вірус, який заражає інші віруси! Але це не станеться без деякої клітинної допомоги.
характеристики вірусу
Віруси можуть бути надзвичайно простими за конструкцією, що складаються з нуклеїнової кислоти, оточеної білковою оболонкою, відомою як капсид. Капсид складається з більш дрібних білкових компонентів називають capsomers. Комбінація капсид+геном називається нуклеокапсидом.
Віруси також можуть володіти додатковими компонентами, причому найбільш поширеним є додатковий мембранозний шар, який оточує нуклеокапсид, званий оболонкою. Оболонка фактично набувається з ядерної або плазматичної мембрани зараженої клітини-господаря, а потім модифікована вірусними білками, званими пепломерами. Деякі віруси містять вірусні ферменти, необхідні для зараження клітини-господаря і закодовані для всередині вірусного генома. Повноцінний вірус, з усіма компонентами, необхідними для інфекції клітин господаря, називають віріоном.
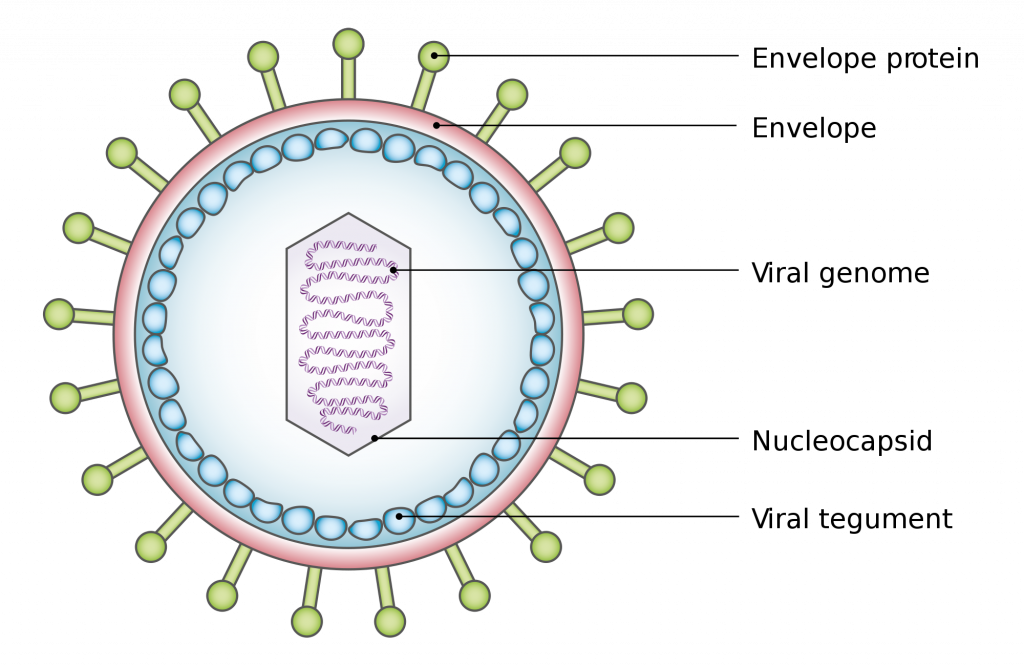
Характеристики вірусу, зображення, створене Беном Тейлором, суспільне надбання, через Вікіпедію
Геном вірусу
У той час як клітини містять двоцепочечной ДНК для свого генома, віруси не обмежуються цією формою. Хоча існують віруси dsDNA, існують також віруси з одноцепочечной ДНК (ssDNA), дволанцюгової РНК (dsRNA) та одноланцюгової РНК (ssRNA). У цій останній категорії SsRNA може або позитивний сенс (+SSRNA, тобто вона може транскрибувати повідомлення, як мРНК), або може бути негативним сенсом (-ssRNA, що вказує на те, що вона доповнює мРНК). Деякі віруси навіть починаються з однієї форми нуклеїнової кислоти в нуклеокапсиді, а потім перетворюють її в іншу форму під час реплікації.
структура вірусу
Вірусні нуклеокапсиди бувають двох основних форм, хоча загальний вигляд вірусу може бути змінений наявністю оболонки, якщо він присутній. Гвинтові віруси мають витягнуту трубкоподібну структуру, причому капсомери розташовані спірально навколо спірального генома. Ікосаедричні віруси мають кулясту форму, з ікосаедричної симетрією, що складається з 20 трикутних граней. Найпростіший ікосаедричний капсид має 3 капсомери на трикутне обличчя, в результаті чого 60 капсомерів для всього вірусу. Деякі віруси не акуратно вписуються ні в одну з двох попередніх категорій, оскільки вони настільки незвичайні за конструкцією або компонентами, тому існує третя категорія, відома як складні віруси. Приклади включають поксвірус з цегляної зовнішністю і складною внутрішньою структурою, а також бактеріофаг з хвостовими волокнами, прикріпленими до ікосаедричної голівці.
Цикл реплікації вірусів
Хоча цикл реплікації вірусів може варіюватися від вірусу до вірусу, існує загальна закономірність, яку можна описати, що складається з п'яти кроків:
- Прикріплення - віріон прикріплюється до правильної клітини-господаря.
- Проникнення або вірусне введення — вірус або вірусна нуклеїнова кислота отримує вхід в клітину.
- Синтез — вірусні білки та копії нуклеїнових кислот виробляються апаратами клітин.
- Асамблея — віруси виробляються з вірусних компонентів.
- Вивільнення — новоутворені віріони вивільняються з клітини.
Вкладення
Поза їх клітини-господаря віруси інертні або метаболічно неактивні. Тому зустріч віріона з відповідною клітиною-господарем є випадковою подією. Сама прив'язка дуже специфічна, між молекулами на зовнішній стороні вірусу і рецепторами на поверхні клітини господаря. Це пояснює специфіку вірусів лише для зараження певних типів клітин або конкретних господарів.
Проникнення або вірусне введення
Багато незагорнуті (або голі) віруси впорскують свою нуклеїнову кислоту в клітину господаря, залишаючи порожній капсид зовні. Цей процес називається проникненням і поширений для бактеріофагів, вірусів, які заражають бактерії. З еукаріотичними вірусами більш імовірно, що весь капсид отримує вхід в клітину, при цьому капсид видаляється в цитоплазмі. Необгорнутий еукаріотичний вірус часто отримує вхід через ендоцитоз, де клітина-господар змушена поглинати капсид, що призводить до ендоцитарного бульбашки. Огорнутий еукаріотичний вірус отримує вхід для свого нуклеокапсиду, коли вірусна оболонка зливається з мембраною клітини господаря, штовхаючи нуклеокапсид повз клітинну мембрану. Якщо весь нуклеокапсид заноситься в клітину, то відбувається процес розкриття, щоб видалити капсид і звільнити вірусний геном.
Синтез
Стадія синтезу значною мірою продиктована типом вірусного генома, оскільки геноми, які відрізняються від генома dsDNA клітини, можуть включати складні вірусні стратегії реплікації генома та синтезу білка. Вірусні специфічні ферменти, такі як РНК-залежні РНК-полімерази, можуть знадобитися для продовження процесу реплікації. Виробництво білка жорстко контролюється, щоб гарантувати, що компоненти виробляються в потрібний час при вірусному розвитку.
Асамблея
Складність вірусної збірки залежить від виробленого вірусу. Найпростіший вірус має капсид, що складається з 3 різних типів білків, які самостійно збираються з невеликими труднощами. Найскладніший вірус складається з понад 60 різних білків, які всі повинні зібратися в певному порядку. Ці віруси часто використовують кілька складальних ліній для створення різних вірусних структур, а потім використовують білки будівельних лісів, щоб організовано зібрати всі вірусні компоненти разом.
Реліз
Більшість вірусів лизують клітину-господаря в кінці реплікації, дозволяючи всім новоутвореним віріонам вивільнятися в навколишнє середовище. Інша можливість, поширена для обволікаючих вірусів, - це бутонізація, коли один вірус вивільняється з клітини одночасно. Клітинна мембрана модифікується шляхом введення вірусних білків, при цьому нуклеокапсид виштовхує через цю модифіковану частину мембрани, дозволяючи їй придбати оболонку.
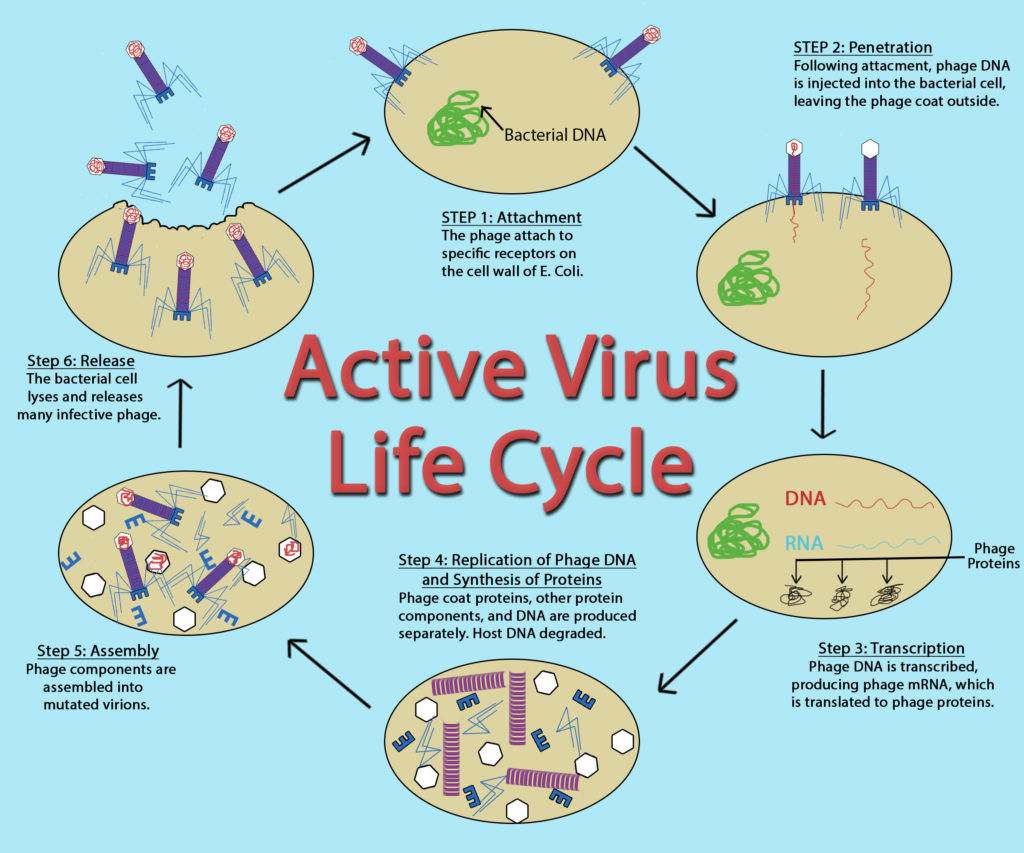
Життєвий цикл активного вірусу Джона Келлогга через OER в Університеті штату Орегон
бактеріофаг
Віруси, що заражають бактерії, відомі як бактеріофаг або фаг. Вірулентний фаг - це той, який завжди лізує клітину-господаря в кінці реплікації, після виконання п'яти етапів реплікації, описаних вище. Це називається літичним циклом реплікації.
Існують також помірний фаг, віруси, які мають два варіанти щодо їх реплікації. Варіант 1 полягає в імітації вірулентного фага, слідуючи п'яти етапам реплікації та лізування клітини-господаря в кінці, що називається літичним циклом. Але помірні фаги відрізняються від вірулентного фага тим, що у них є інший вибір: варіант 2, де вони залишаються всередині клітини-господаря, не руйнуючи її. Цей процес відомий як лізогенез або лізогенний цикл реплікації.
Фаг, який використовує лізогенез, все ще проходить перші два етапи типового циклу реплікації, приєднання та проникнення. Після того, як вірусна ДНК була вставлена в клітину, вона інтегрується з ДНК господаря, утворюючи профаг. Заражену бактерію називають лізогенної або лізогенної бактерією. У такому стані вірус користується стабільними відносинами зі своїм господарем, де він не перешкоджає метаболізму або розмноженню клітин господаря. Клітина господаря користується імунітетом від повторного зараження від того ж вірусу.
Вплив клітини-господаря стресових станів (тобто УФ-опромінення) викликає індукцію, де вірусна ДНК висікає з ДНК клітини-господаря. Ця подія запускає решту етапів літичного циклу, синтезу, дозрівання та вивільнення, що призводить до лізису клітини-господаря та вивільнення новоутворених віріонів.
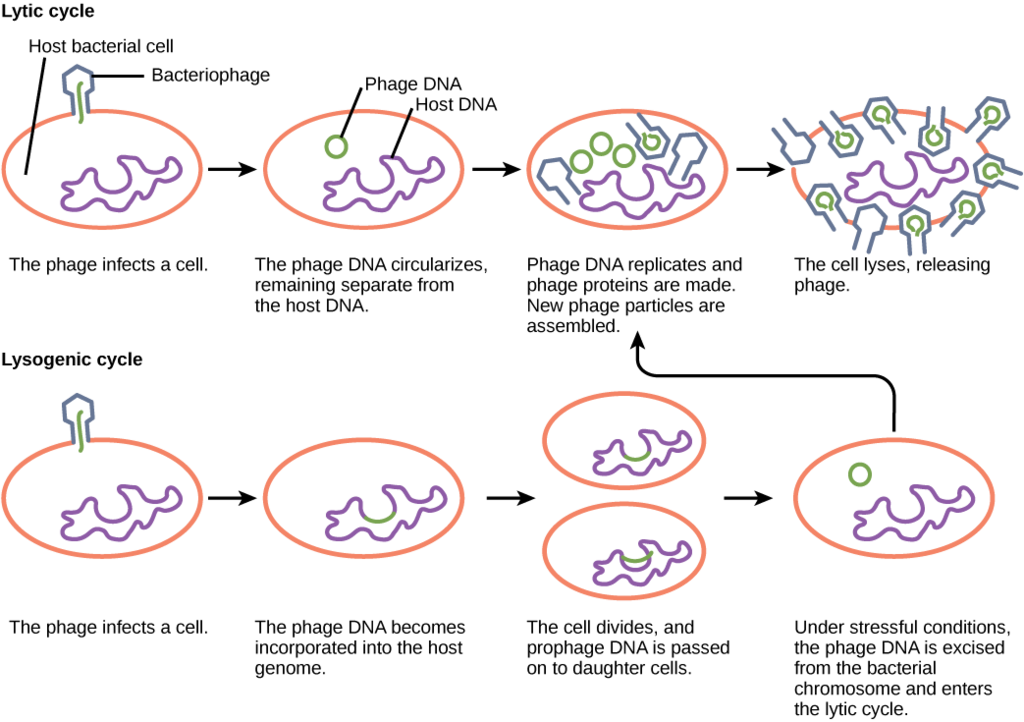
Літичний цикл проти лізогенного циклу реплікації. OpenStax, вірусні інфекції та хости. OpenStax CNX. 11 квіт 2013 р. http://cnx.org/contents/7cbd15ad-5bff-4678-a99f-85fd579e070c@3.
Отже, що ж диктує тип реплікації, який буде використовуватися помірним фагом? Якщо навколо багато клітин господаря, цілком ймовірно, що помірний фаг буде брати участь у літичному циклі реплікації, що призведе до великого збільшення виробництва вірусів. Якщо клітини-господаря дефіцитні, помірний фаг частіше вступає в лізогенез, що дозволяє вірусне виживання, поки кількість клітин господаря не збільшиться. Те ж саме вірно, якщо кількість фагів у середовищі значно перевищує кількість клітин господаря, оскільки лізогенія дозволила б відскочити кількість клітин господаря, забезпечуючи довгострокове виживання вірусів.
Лізогени також можуть відчувати користь від лізогенезу, оскільки це може призвести до лізогенної конверсії, ситуації, коли розвиток пророцтва призводить до зміни фенотипу господаря. Одним з кращих прикладів цього є бактерія Corynebacterium diphtheriae, збудника дифтерії. Дифтерійний токсин, який викликає захворювання, кодується в геномі фагів, тому дифтерію викликають лише C. diphtheriae lysogens.
Віруси еукаріотів
Еукаріотичні віруси можуть викликати один з чотирьох різних результатів для їх клітини-господаря. Найбільш поширеним результатом є лізис клітин господаря, що виникає в результаті вірулентної інфекції (по суті, літичний цикл реплікації спостерігається у фагів). Деякі віруси можуть викликати приховану інфекцію, співіснуючи мирно зі своїми клітинами-господарями роками (так само, як помірний фаг під час лізогенезу). Деякі огорнуті еукаріотичні віруси також можуть виділятися по одному з зараженої клітини-господаря у вигляді процесу бутонізації, викликаючи стійку інфекцію. Нарешті, певні еукаріотичні віруси можуть спричинити перетворення клітини-господаря в злоякісну або ракову клітину, механізм, відомий як трансформація.
Віруси і рак
Існує багато різних причин раку, або нерегульованого росту і розмноження клітин. Деякі відомі причини включають вплив певних хімічних речовин або ультрафіолетового світла. Існують також певні віруси, які мають відомі пов'язані з розвитком раку. Такі віруси відносять до онковірусів. Онковіруси можуть викликати рак, виробляючи білки, які зв'язуються з білками господаря, відомими як білки-супресори пухлини, які функціонують для регулювання росту клітин та ініціювання запрограмованої загибелі клітин, якщо це необхідно. Якщо білки-супресори пухлини інактивуються вірусними білками, то клітини виростають з-під контролю, що призводить до розвитку пухлин і метастазування, де клітини поширюються по всьому організму.
Ключові слова
вірус, облігатний внутрішньоклітинний паразит, капсид, бактеріофаг, капсомер, нуклеокапсид, оболонка, пепломер, віріон, dsDNA, SSDNA, дСРНК, +SSRNA, спіральні віруси, ікосаедричні віруси, складні віруси, приєднання, проникнення, вірусне введення, синтез, збірка, вивільнення, голий вірус, ендоцитоз, бутонізація, бактеріофаг, фаг, вірулентний фаг, літичний цикл, помірний фаг, лізогенез, лізогенний цикл, профаг, лізоген, лізогенна бактерія, індукція, лізогенна конверсія, вірулентна інфекція, латентна інфекція, персистуюча інфекція, трансформація, онковірус, білки-супресори пухлини.
Основні питання/цілі
- Які загальні властивості вірусу?
- Який розмірний ряд вірусів? Як вони порівнюють за розмірами з бактеріями?
- Яке загальне будова вірусів? Які існують різні компоненти?
- Які вірусні форми існують?
- Як конверти і ферменти відносяться до вірусів?
- Які типи вірусних геномів існують?
- Які етапи розмноження вірусів? Що відбувається на кожному кроці? Чим бактеріальні/архейальні віруси відрізняються від вірусів еукаріотичних щодо розмноження?
- Які 2 типи вірусної інфекції виявляються в бактерії/археї? Які конкретні терміни пов'язані з вірусною інфекцією бактеріальних/архейних клітин?
- Які 4 типи вірусної інфекції виявляються у еукаріотів?
- Як деякі віруси викликають рак?
Дослідницькі питання (НЕОБОВ'ЯЗКОВО)
- Яка найбільша бактерія або архея коли-небудь виявлена? Який найменший еукаріот коли-небудь виявлений?