9: Еволюційність та пластичність
- Page ID
- 3399

Еволютивність виникла як зв'язок між еволюцією на рівні населення та макроеволюцією. Хоча обидва ці процеси принципово однакові (обидва відбуваються через мутацію, дрейф та відбір), вони відбуваються в таких різних масштабах, що ми зазвичай використовуємо дуже різні інструменти та мислення для вивчення кожного з них. Наприклад, еволюція на рівні населення часто моделюється рівняннями, пов'язаними з Харді-Вайнбергом, вивчаючи придатність та поширеність певних алелів. З іншого боку, ми зазвичай вивчаємо макроеволюційні зміни, порівнюючи різні види за допомогою викопного запису та молекулярної генетики. Питання в тому, як ми можемо перейти звідси туди? Як ці процеси на мікрорівні в кінцевому підсумку призводять до шаблонів макрорівня? Evolvability є одним з корисних способів наблизитися до цього. Населення є «еволюціонуючим», якщо воно може впоратися зі зміною середовища за допомогою адаптації. Тобто, якщо генетичні мутації в популяції мають хороші шанси на підвищення фітнесу.
Хоча на поверхневому рівні це здається спрощеним, ми можемо слідувати за ним до більш глибоких рівнів, щоб побачити, як еволюція населення впливає на процеси розвитку, еволюція яких лежить в основі макроеволюційних змін. Герхарт і Кіршнер вирішують це у своїй роботі Теорія полегшеної варіації. У цій роботі вони визначають основні шляхи, за допомогою яких генетичні зміни розвитку можуть «полегшити» (або допомогти збільшити) фенотипічні варіації без шкоди для придатності (тобто вони підвищують еволюцію).
Ці маршрути, які називаються слабкими регуляторними зв'язками, дослідницькими процесами та розділенням, підсумовуються нижче:
Здатність брати участь у слабкому регулювальному зв'язку
Зв'язок відбувається між процесами, які пов'язані один з одним або з однаковими конкретними умовами. Наприклад, коли вам холодно, ви тремтіте і отримуєте мурашки по шкірі. Ці два процеси пов'язані один з одним через стан (холод) а також через симпатичну нервову систему. Так само процеси розвитку можуть бути пов'язані через спільні регуляторні мережі генів та/або через спільні регуляторні компоненти (наприклад, гени, які виражені у відповідь на той же фактор транскрипції, або шляхи трансдукції сигналів, які реагують на ту саму сигнальну молекулу).
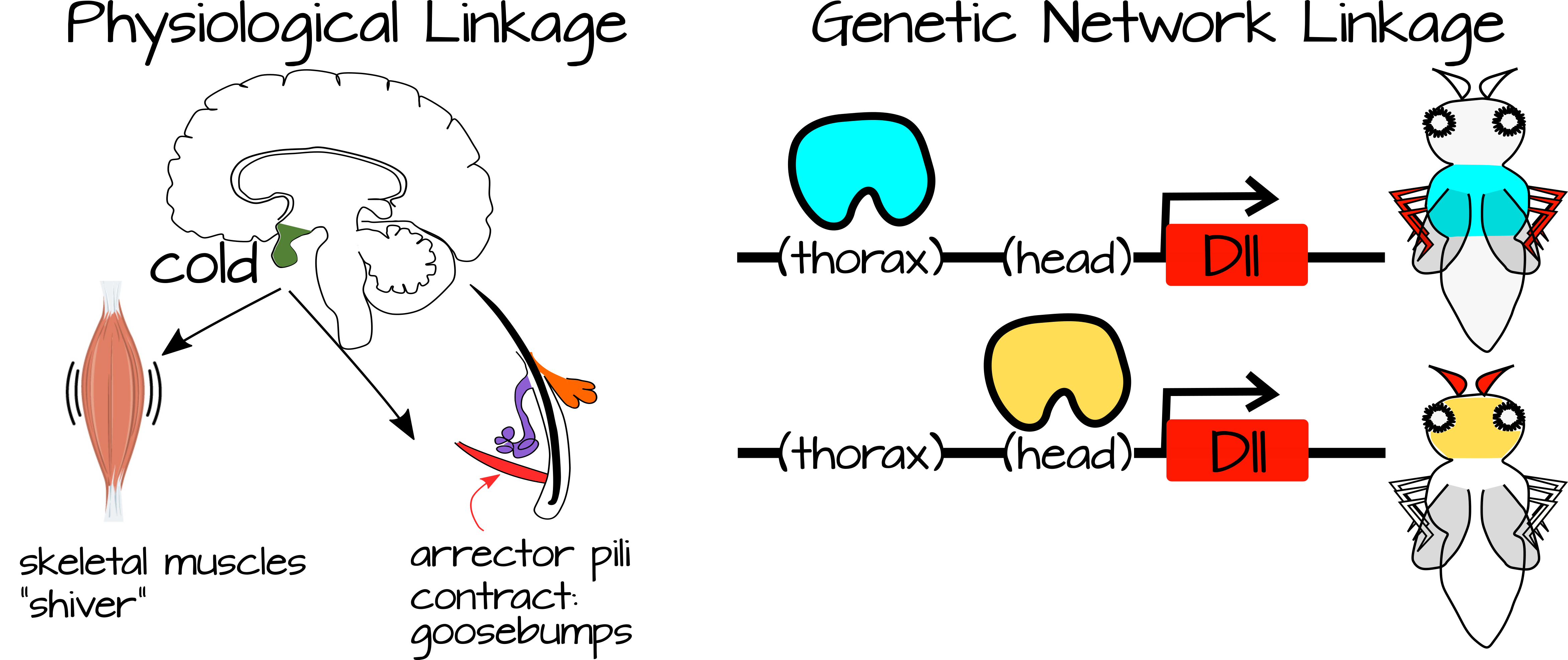
Слабкий регуляторний зв'язок стосується пов'язаних процесів, які регулюються простими входами, які не надають багато інформації процесам. Ми бачили приклади цього в звичках високоефективних сигнальних шляхів, де фактор транскрипції вивільняє репресії на набір генів нижче за течією. Ці гени самоактивуються за допомогою місцевих активаторів, як тільки вивільняється інгібування, припускаючи, що потенційно може бути кілька способів їх деінгібування. Це вважається слабким регуляторним зв'язком, оскільки регулювання фактором транскрипції є «слабким». Коли локальні активатори присутні, за замовчуванням для процесу є «on». Фактор транскрипції потрібен лише для звільнення гальмування. Після звільнення цього гальмування може статися каскад експресії генів, що призводить до складного процесу з великими наслідками розвитку або фізіологічними. Таким чином складний процес може бути вимкнений або включений в нових місцях з невеликими нормативними змінами. Наприклад, експресія фактора транскрипції може бути повернута вгору або вниз, або інші білки можуть діяти, щоб змінити здатність фактора транскрипції зв'язуватися зі своєю цільовою ДНК.
Загалом, слабкий регуляторний зв'язок виникає, коли простий сигнал може викликати кілька складних процесів залежно від контексту стільникового зв'язку. Герхарт і Кіршнер відзначають, що це збільшує пластичність системи, оскільки невеликі зміни регуляторних факторів (наприклад, факторів транскрипції) можуть змінити функціональний вихід складної системи. Сама складна система багато в чому саморегульована, що вимагає лише спускового механізму від регуляторних факторів. Таким чином, розвиток може відбуватися повільніше або швидше за певних умов, вибір між двома станами тканини (наприклад, типом гонади) може бути змінений факторами навколишнього середовища тощо.
Розвідувальні процеси
Дослідницькі процеси - це пошук та пошук процесів, таких як проходження хімічного градієнта до точкового джерела. Ми бачимо цей тип процесу в розвитку, коли вивчаємо, як утворюються аксони під час нейрогенезу і як виникає судинна система для живлення органів і тканин. Ми бачимо це у дорослих в адаптивній імунній системі хребетних та харчовій поведінці в хімічно керованих організмах, таких як мурахи. Ми говоримо, що цей тип процесу демонструє надійність, оскільки він швидко адаптується до місцевих екологічних змін. Наприклад, при суворому малюнку сітки XY судинної системи ріст протягом часу розвитку повинен бути під суворим контролем, а дисперсія розміру/форми в органах не допускається. Малі кровоносні судини, однак, не ростуть через систему координат, скоріше вони ростуть на основі системи «попиту і пропозиції» з виростами в області низького кисню. Клітини виділяють білковий сигнал (VEGF), коли вони мають низький вміст кисню, сприяючи зростанню кровоносних судин до них 2.
Дослідницькі процеси також адаптуються протягом еволюційного часу, оскільки вони дозволяють відхилення розмірів та форми. Це важливо для популяцій, де особини можуть відрізнятися за розміром або формою. Але це також важливо протягом більш тривалих періодів еволюційного часу, оскільки ці гнучкі процеси малюнка сформують корисну систему, засновану на їх простому наборі правил зростання. Наприклад, пересаджені диски ніг у комах іннервуються, як і позаматкові кінцівки у пташенячих ембріонів 3,4. Таким чином, дослідницькі процеси полегшують еволюцію, допомагаючи побудувати життєздатне тіло, коли інші морфологічні компоненти перетворилися на більш придатні конформації.
Відділення
У рослин здатність відокремлювати світлові реакції від циклу фотосинтезу Кальвіна або тимчасово (як у рослин CAM), або просторово (як у рослин С4), призвела до неймовірного успіху та різноманітності в цих типах рослин, оскільки вони більше не залежать від збереження своїх продихів відкритими під час найспекотніших і найсухіші (але і найсонячніші) частини дня. У наших власних клітині ми можемо побачити компартменталізацію наших мітохондрій, причому міжмембранний простір приблизно в 10 разів кислотніше, ніж просвіт. Це дозволяє концентрувати протонну рушійну силу, а також захищає рН решти клітини. Наші клітини також фізично відокремлюють клітинну сигналізацію від транскрипційної регуляції - дозволяючи модифікувати сигнальні шляхи залежно від історії клітин (розвитку) та навколишнього середовища.
Коли одне відділення організму (або клітини) може діяти напівнезалежно від іншого, ми очікуємо більш високих варіацій. Тобто кожен відсік може запустити свою програму розвитку за допомогою активації та гальмування конкретних GRN, не порушуючи розвитку інших відсіків. Таким чином, зміни експресії генів, які зачіпають лише один або кілька відсіків, можуть бути обмежені в дії. Таким чином, еволюція може впливати на відсіки окремо, якщо це зміни вираження, такі як кількість стенограми, ізоформи білка або посттрансляційна модифікація. Мутації, що кодують білок, з іншого боку, потенційно можуть діяти на багатьох відділеннях одночасно. Еволюція потенційно може впливати на окремі відсіки через зміни експресії генів, тоді як дослідницькі процеси можуть підтримувати надійність сигнальних та поживних шляхів у всьому тілі.
Еволютивність у тварин залежить від двох конкуруючих типів розвитку:
- Процеси стійкі до еволюційних змін. До них відносяться дослідницькі процеси, які використовують зовнішні сигнали для пошуку найбільш ефективної або найбільш ефективної морфології, що призводить до збереження процесу. Але вони також можуть включати процеси, які зберігаються протягом тривалих еволюційних періодів часу через фізичні або генетичні обмеження.
- Змінні процеси. До них відносяться процеси, які змінюються з часом, оскільки вони або розділені, або кооптизовані на інші надійні програми розвитку. Компартментація (процес, стійкий до змін) дозволяє фізично розділити процеси розвитку, а слабкий регуляторний зв'язок дозволяє коопціювати процеси до нових часів і місць (наприклад, відсіків).
Таким чином еволюційні зміни відбуваються на тлі робастних процесів. Деякі з цих робастних процесів є податливими у часі розвитку (наприклад, васкуляризація), тоді як інші є обмеженнями - обмежуючи те, що можливо в певній еволюційній лінії. Один із прикладів, який ми вже розглядали, - еволюція чотириногих кінцівок. Хоча існують фізичні та генетичні обмеження на це обмеження кількості ущільнень кісток на секцію розвитку, ці обмеження також створюють відсіки - стилопод, зеугопод, і аутопод, а також 5 цифр. Зміна розміру та адаптація до відсіків підтримується надійними дослідницькими процесами, пов'язуючи розвиток кісток з підтримкою мускулатури, судинної системи та іннервації.
Подальше читання
Глибока гомологія (генетичні обмеження): https://www.sdbonline.org/sites/fly/lewhelddeep/deephomology.htm
Бауплани (обмеження розвитку): https://evolution-outreach.springeropen.com/articles/10.1007/s12052-012-0424-z
Генетика модулюється навколишнім середовищем, ця модуляція може бути короткочасною (наприклад, отримання засмаги) або довгоживучою (наприклад, отримання інсулінорезистентного діабету). Він може охоплювати навіть покоління (ощадливий епігенотип). Як виникає ця «пластичність» в генетиці і як це змінює те, як працює еволюція?
http://science.sciencemag.org/content/357/6350/eaan0221.long: Якщо ви зробите свій генотип 23andme, він лише розповість вам, який ваш ризик для певних захворювань та розладів. Чому ми не можемо бути впевнені?
Посилання
- «Теорія полегшеної варіації» 2007, PNAS, Джон Герхарт та Марк Кіршнер, https://doi.org/10.1073/pnas.0701035104
- Переглянуто в «Гіпоксия-індукований ангіогенез: добро і зло», 2011, Гени та рак, Брайан Крок, Ніколас Скулі. і М. Селеста Саймон doi: 10.1177/1947601911423654
- «Аксональні проекції з пересаджених позаматкових ніг у комахи», 1985, Журнал порівняльної неврології, П.Сівасубраманіан D.R. Nässel, doi.org/10.1002/cne.902390211
- «Іннервація FGF-індукованих додаткових кінцівок у пташенячого ембріона» 2003, Журнал анатомії, BW Turney, AM Роуан-Халл, і Дж.М. Браун doi: 10.1046/j.1469-7580.2003.00131.x
- «Ангіогенез: адаптивна проблема динамічного біологічного малюнка», 2013, Обчислювальна біологія PLOS, Тімоті В. Сікомб, Джонатан П. Альбердінг, Річард Хсу, Марк Дьюірст, Аксель Прайс https://doi.org/10.1371/journal.pcbi.1002983
- «Нейронна міграція та ламінування в сітківці хребетних», 2018, Межі в неврології, Рана Аміні, Маурісіо Роча-Мартінс та Карен Норден. doi: 10.3389/fnins.2017.00742
Мініатюра — це графік постійної варіації популяції (10 000 поліморфних сайтів з людської хромосоми 1) Грема Купа, опублікований на Вікісховищі за ліцензією CC BY 3.0.