13: Генетичний код
- Page ID
- 8020

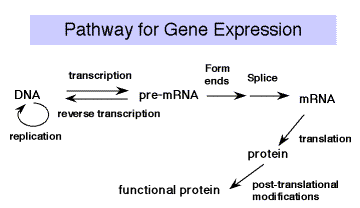
Після завершення транскрипції та обробки РРНК, ТРНК та SNRNA РНК готові до використання в клітині - зібрані в рибосоми або SNRNP і використовуються для сплайсингу та синтезу білка. Але зріла мРНК ще не функціональна для клітини. Його необхідно перевести в закодований білок. Правила перекладу з «мови» нуклеїнових кислот на білки - це генетичний код.
Вступ
Експерименти з тестуванням ефектів мутацій зрушення кадру показали, що видалення або додавання 1 або 2 нуклеотидів викликало втрату функції, тоді як видалення або додавання нуклеотидів 3 дозволило зберегти значну функцію. Це продемонструвало, що одиницею кодування є 3 нуклеотиди. Нуклеотидний триплет, який кодує амінокислоту, називається кодоном. Кожна група з трьох нуклеотидів кодує по одній амінокислоті. Оскільки існує 64 комбінації з 4 нуклеотидів, взятих по три за раз і всього 20 амінокислот, код є виродженим (більше одного кодону на амінокислоту, в більшості випадків). Молекулою адаптера для перекладу є тРНК. Заряджена тРНК має амінокислоту на одному кінці, а на іншому - антикодон для узгодження кодону в мРНК; тобто. він «говорить мовою» нуклеїнових кислот на одному кінці і «мовою» білків на іншому кінці. Машиною синтезу білків під керівництвом шаблонної мРНК є рибосома.
Малюнок 3.4.1 ТРНК служать адаптером для перекладу з нуклеїнової кислоти в білок
А. розмір кодону: 3 нуклеотиди
1. Три - мінімальна кількість нуклеотидів на кодон, необхідне для кодування 20 амінокислот.
а. 20 амінокислот кодуються комбінаціями з 4 нуклеотидів
б Якби кодон був двома нуклеотидами, набір всіх комбінацій міг кодувати тільки
4х4 = 16 амінокислот.
с. з трьома нуклеотидами набір всіх комбінацій може кодувати
4x4x4 = 64 амінокислоти (тобто 64 різних комбінації з чотирьох нуклеотидів, взятих по три за раз).
2. Результати комбінацій мутацій зсуву кадрів показують, що код знаходиться в трійках. Мутації, що змінюють довжину, які додають або видаляють один або два нуклеотиди, мають серйозний дефектний фенотип (вони змінюють кадр зчитування, тому вся послідовність амінокислот після мутації змінюється.). Але ті, які додають або видаляють три нуклеотиди, мало або зовсім не впливають. В останньому випадку кадр зчитування підтримується, з введенням або видаленням амінокислоти на одній ділянці. Комбінації трьох різних одиничних нуклеотидних делецій (або вставок), кожна з яких має фенотип втрати функції окремо, можуть відновити істотну функцію гена. Кадр читання wild типу відновлюється після 3-го видалення (або вставки).
Б. експерименти з розшифровки коду
1. Було розроблено кілька різних безклітинних систем, які каталізують синтез білка. Ця здатність здійснювати переклад in vitro була одним із технічних досягнень, необхідних для того, щоб дозволити дослідникам визначити генетичний код.
а. ретикулоцити ссавців (кролика): рибосоми активно виробляють багато глобіну.
б. екстракти зародків пшениці
c Бактеріальні екстракти
2. Здатність синтезувати випадкові полінуклеотиди була ще однією ключовою розробкою, яка дозволила експериментам розшифрувати код. S. Ochoa виділив фермент полінуклеотидфосфорилазу і показав, що він здатний зв'язувати нуклеозиди ді фосфати (НДП) в полімери НМП (РНК) в оборотній реакції.
nNDP n + NPi
Фізіологічна функція полінуклеотидної фосфорилази полягає в каталізації зворотної реакції, яка використовується при деградації РНК. Однак у безклітинній системі пряма реакція дуже корисна для виготовлення випадкових РНК полімерів.
3. Гомополімери програмують синтез специфічних гомо-поліпептидів (Nirenberg and Matthei, 1961).
а Якщо ви надаєте лише UDP як субстрат для полінуклеотидної фосфорилази, продукт буде гомополімерним полі (U).
б Додавання полі (U) до системи перекладу in vitro (наприклад, лізатів кишкової палички) призводить до нещодавно синтезованого поліпептиду, який є полімером поліфенілаланіну.
c. таким чином UUUU кодує Phe.
d Аналогічно, полі (А) запрограмований синтез Poly‑LYS; AAA кодує Lys.
Полі (C) запрограмований синтез Poly‑Pro; CCC кодує Pro.
Полі (G) запрограмований синтез Poly‑GLY; GGG кодує Gly.
4. Використання змішаних сополімерів
а Якщо два НДФ змішуються у відомому співвідношенні, полінуклеотидна фосфорилаза утворює змішаний сополімер, в який нуклеотид включений з частотою, пропорційною його присутності в вихідній суміші.
б Наприклад, розглянемо суміш 5:1 A: C Фермент буде використовувати ADP 5/6 часу, і CDP 1/6 часу. Прикладом можливого продукту є:
ААКААААААААААААААААААААААААААА
|
Склад |
Число |
Імовірність |
Відносна частота |
|---|---|---|---|
|
3 А |
1 |
0,578 |
1.0 |
|
2 А, 1 СМ |
3 |
3 х 0,116 |
3 х 0,20 |
|
1 А, 2 СМ |
3 |
3 х 0,023 |
3 х 0,04 |
|
3 СМ |
1 |
0,005 |
0,01 |
c Таким чином, частота, що AAA буде відбуватися в сополімері, дорівнює
(5/6) (5/6) (5/6) = 0,578.
Це буде найбільш часто зустрічається кодон, і його можна нормалізувати до 1,0 (0,578/0,578 = 1,0)
d Частота, яку буде відбуватися кодон з 2 А і 1 С, дорівнює
(5/6) (5/6) (1/6) = 0,116.
Є три способи мати 2 А і 1 С, тобто AAC, ACA і CAA. Так частота виникнення всіх кодонів A2C дорівнює 3 х 0,116.
Нормалізуючи на ААА, що має відносну частоту 1,0, частота кодонів A2C дорівнює 3 х (0,116/0,578) = 3 х 0,2.
е. аналогічна логіка показує, що очікувана частота кодонів AC2 становить 3 х 0,04, а очікувана частота CCC дорівнює 0,01.
|
Радіоактивний |
Осадова cpm |
Спостерігається |
Теоретичний |
|||||
|---|---|---|---|---|---|---|---|---|
|
амінокислоти |
- шаблон |
+ шаблон |
включення |
включення |
||||
|
Лізин |
60 |
4615 |
100.0 |
100 |
||||
|
Треонін |
44 |
1250 |
26.5 |
24 |
||||
|
аспарагін |
47 |
1146 |
24.2 |
20 |
||||
|
Глютамин |
39 |
1117 |
23.7 |
20 |
||||
|
Пролін |
14 |
342 |
7.2 |
4.8 |
||||
|
Гістидин |
282 |
576 |
6.5 |
4 |
||||
Ці дані взяті з Speyer et al. (1963) Симпозіум Cold Spring Harbor з кількісної біології, 28:559. Теоретичне включення - це очікуване значення, враховуючи генетичний код, як він був згодом визначений.
f Коли ця суміш змішаних сополімерів використовується для програмування перекладу in vitro, Lys включається найчастіше, що може бути виражено як 100. Це підтверджує, що AAA кодує Lys.
г Відносно Lys включення як 100, Thr, Asn і Gln включені зі значеннями від 24 до 26, дуже близькі до очікування для амінокислот, кодованих одним з кодонів A2C. Однак ці дані не показують, який з кодонів A2C кодує кожну конкретну амінокислоту. Тепер ми знаємо, що ACA кодує Thr, AAC кодує Asn, а CAA кодує Gln.
h Pro і His включені зі значеннями 6 і 7, що близьке до очікуваних 4 для амінокислот, кодованих кодонами AC2. Наприклад, CCA кодує Pro, CAC кодує його. ACC кодує Thr, але це включення затьмарюється «26.5" одиницями реєстрації в ACA. Або, точніше, «26.5» @ 20 (ACA) + 4 (АСС) для Thr.
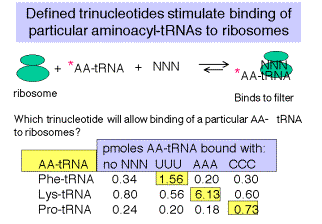
5. Визначені тринуклеотидні кодони стимулюють зв'язування аміноацил‑ТРНК з рибосомами
а. при високих концентраціях катіонів Mg 2+ нормальний механізм ініціювання, що вимагає F‑met‑TRNAF, може бути змінений, а визначені тринуклеотиди можуть бути використані для прямого зв'язування окремих мічених аміноацил‑ТРНК з рибосомами.
б. Наприклад, якщо рибосоми змішуються з UUU та радіомаркованим PHE-trNaphe, за цих умов утворюється потрійний комплекс, який буде прилипати до нітроцелюлози («аналіз Millipore», названий на честь виробника нітроцелюлози).
c Потім можна перевірити всі можливі комбінації триплетних нуклеотидів.
Малюнок 3.4.2. Дані Ніренберга та Ледера (1964) Наука 145:1399.
6. Повторювана послідовність синтетичних полінуклеотидів (Хорана)
a. чергуються сополімери: наприклад (UC) в програмах включення Ser і Leu. Так УКУ і CUC кодують Сер і Лей, але не можуть сказати, що є що. Але в поєднанні з іншими даними, наприклад випадковими змішаними сополімерами в розділі 4 вище, можна зробити деякі остаточні визначення. Така подальша робота показала, що УКУ кодує Ser і CUC кодує лей.
b. Полі (AUG) програми включення Poly‑MET та Poly‑ASP при високих концентраціях Mg. AUG кодує Met, UGA - це зупинка, тому ГУА повинен кодувати Asp.
C. Генетичний код
Шляхом складання спостережень з експериментів, таких як описані в попередньому розділі, була визначена кодувальна здатність кожної групи з 3 нуклеотидів. Це іменується генетичним кодом. Вона зведена в табл. 3.4.4. Це говорить нам про те, як клітина перекладається з «мови» нуклеїнових кислот (полімерів нуклеотидів) на білки (полімери амінокислот).
Таблиця 3.4.4. Генетичний код
Положення в Кодоні
|
1-й |
2-й. |
3-й |
||||||||||||||
|
У. |
С. |
А. |
Г. |
|||||||||||||
|
У |
УУУ |
Фе |
УКУ |
Сер |
УАУ |
Тир |
УГУ |
Cys |
У |
|||||||
|
УУЦ |
Фе |
UCC |
Сер |
УАК |
Тир |
УГК |
Cys |
C |
||||||||
|
УУА |
Лей |
УКА |
Сер |
УАА |
Термін |
ГУГА |
Термін |
A |
||||||||
|
УУГ |
Лей |
УСГ |
Сер |
АГ |
Термін |
УГГІ |
Трп |
Г |
||||||||
|
C |
КУУ |
Лей |
CCU |
Про |
КАУ |
Його |
CGU |
Арг |
У |
|||||||
|
КУС |
Лей |
CCC |
Про |
CAC |
Його |
CGC |
Арг |
C |
||||||||
|
КУА |
Лей |
CCA |
Про |
CAA |
Глен |
CGA |
Арг |
A |
||||||||
|
КУГ |
Лей |
ККГ |
Про |
КЛІТКА |
Глен |
CGG |
Арг |
Г |
||||||||
|
A |
АУУ |
Іль |
АКУ |
Thr |
ААУ |
Асн |
АГУ |
Сер |
У |
|||||||
|
АУК |
Іль |
АКК |
Thr |
AAC |
Асн |
АГЦ |
Сер |
C |
||||||||
|
АУА |
Іль |
АКА |
Thr |
ААА |
Лис |
АГА |
Арг |
A |
||||||||
|
СЕРПЕНЬ* |
Зустріли |
АКГ |
Thr |
ААГ |
Лис |
АГГ |
Арг |
Г |
||||||||
|
Г |
ГУУ |
Вал |
ГКУ |
Ала |
ГАУ |
Жерех |
ГГУ |
Гли |
У |
|||||||
|
ГУК |
Вал |
GCC |
Ала |
ГАК |
Жерех |
GGC |
Гли |
C |
||||||||
|
ГУА |
Вал |
GCA |
Ала |
ГАА |
Глю |
ГГА |
Гли |
A |
||||||||
|
ГУГ* |
Вал |
GCG |
Ала |
КЛЯП |
Глю |
ГГГ |
Гли |
Г |
||||||||
* Іноді в якості ініціатора використовуються кодони.
2. Із загальної кількості 64 кодонів 61 кодує амінокислоти і 3 вказують припинення перекладу.
3. Виродження
Виродження генетичного коду відноситься до того, що більшість амінокислот уточнюється більш ніж одним кодоном. Виняток становлять метіонін (AUG) і триптофан (UGG). Виродження зустрічається в першу чергу третя позиція. Отже, одиночні нуклеотидні заміни на третій позиції можуть не привести до зміни закодованої амінокислоти. Вони називаються безшумними або синонімічними замінами нуклеотидів і не змінюють закодований білок. Про це більш детально розглянуто нижче.
Патерн виродження дозволяє організувати кодони на "сім'ї" та "пари». У 9 групах кодонів нуклеотидів на перших двох позиціях достатньо, щоб вказати унікальну амінокислоту, а будь-який нуклеотид (скорочено N) на третій позиції кодує ту саму амінокислоту. До них належать 9 кодонових «сімейств». Прикладом може служити ACN кодування треонін.
Існує 13 кодонових «пар», в яких нуклеотидів на перших двох позиціях досить вказати дві амінокислоти. Нуклеотид пурину (R) у третій позиції визначає одну амінокислоту, тоді як нуклеотид піримідину (Y) у третій позиції визначає іншу амінокислоту.
Ці приклади додають до більш ніж 20 (кількість амінокислот), оскільки лейцин (кодується UUR та CUN), серин (кодується UCN та AGY) та аргінін (кодується CGN та AGR) кодуються як сімейством кодонів, так і кодоновою парою. Кодони UAR, що вказують на припинення перекладу, зараховувалися як кодонова пара. Три кодони, що кодують ізолейцин (AUU, AUC та AUA), знаходяться на півдорозі між сімейством кодонів та парою кодонів.
4. Хімічно схожі амінокислоти часто мають схожі кодони.
5. Основним кодоном, що визначає ініціацію перекладу, є AUG
Бактерії також можуть використовувати GUG або UUG, і дуже рідко AUU і, можливо, CUG. Використовуючи дані 4288 генів, ідентифікованих повною послідовністю геному кишкової палички, була визначена наступна частота використання кодонів в ініціації:
- AUG використовується для 3542 генів.
- GUG використовується для 612 генів.
- УУГ використовується для 130 генів.
- AUU використовується для 1 гена.
- CUG може бути використаний для 1 гена.
Незалежно від того, який кодон використовується для ініціації, перша амінокислота, включена під час трансляції, - це F-мет у бактерій.
6. Три кодони вказують припинення перекладу: UAA, UAG, UGA.
З цих трьох кодонів УАА використовується найчастіше в кишковій паличці, а потім УГА. UAG використовується набагато рідше.
- УАА використовується для 2705 генів.
- УГА використовується для 1257 генів.
- УАГ використовується для 326 генів.
7. Генетичний код практично універсальний
У рідкісних винятках з цього правила відмінності від генетичного коду досить малі. Наприклад, одним винятком є РНК з мітохондріальної ДНК, де і UGG, і UGA кодують Trp.
D. диференціальне використання кодонів
1. Різні види мають різні моделі використання кодонів: Наприклад, можна використовувати 5' UUA для кодування Leu 90% часу (визначається нуклеотидними послідовностями багатьох генів). Він може ніколи не використовувати CUR, а комбінація UUG плюс CUY може становити 10% кодонів.
2. Велика кількість тРНК корелює з використанням кодонів у природних мРНК: У цьому прикладі TrnAleu з 3' AAU на антикодоні буде найбільш поширеним.
3. Схема використання кодонів може бути предиктором рівня експресії гена: Загалом, більш виражені гени, як правило, використовують кодони, які часто використовуються в генах в решті частини генома. Це було кількісно оцінено як «індекс адаптації кодонів». Таким чином, аналізуючи повні геноми, раніше невідомий ген, профіль використання кодону якого відповідає бажаному використанню кодону для організму, високо оцінював би індекс адаптації кодону, і можна було б припустити, що це високоекспресований ген. Так само той, хто має низький бал за індексом, може кодувати білок з низькою кількістю.
Спостереження за геном із схемою використання кодону, яка суттєво відрізняється від решти генома, вказує на те, що цей ген, можливо, потрапив у геном шляхом горизонтального перенесення від іншого виду.
4. Переважне використання кодону є корисним міркуванням у «зворотній генетиці»: Якщо ви знаєте навіть часткову послідовність амінокислот для білка і хочете виділити ген для нього, сімейство послідовностей мРНК, які можуть кодувати цю послідовність амінокислот, можна легко визначити. Через виродження в коді це сімейство послідовностей може бути дуже великим. Оскільки ці послідовності, ймовірно, будуть використовувати як зонди гібридизації або як праймери ПЛР, чим більше сімейство можливих послідовностей, тим більша ймовірність того, що можна отримати гібридизацію до цільової послідовності, яка відрізняється від бажаної. Таким чином, хочеться обмежити кількість можливих послідовностей, і, посилаючись на таблицю переваг кодонів (припускаючи, що вони відомі для організму, що цікавить), то можна використовувати бажані кодони, а не всі можливі кодони. Це обмежує кількість послідовностей, які потрібно зробити як зонди гібридизації або праймери.
Е. коливання в антикодоні
Ця гнучкість в положенні «коливання» дозволяє деяким ТРНК з'єднуватися з двома або трьома кодонами, тим самим зменшуючи кількість ТРНК, необхідних для перекладу. Наступні правила «коливання» означають, що 61 кодон (для 20 амінокислот) можна прочитати лише 31 антикодон (або 31 тРНК).
Окрім звичайних базових пар, можна мати пари G‑U, а I у 1-й позиції антикодону можна з'єднати з U, C або A (правила коливання).
5' основа антикодону = 3' база кодону =
перша позиція в третю позицію тРНК в мРНК
С Г
А У
U A або G
G C або U
I U, C або A

F. Види мутацій
Базові заміни
Це вже було висвітлено в частині другій, ремонт ДНК. Як нагадування, існує два типи базових замін.
- Переходи: замінники пурину пурину або замінники піримідину іншого піримідину. Той же клас нуклеотидів залишається. Прикладами є A, що замінює G або C, замінюючи T.
- Трансверсії: замінники пурину піримідину або замінники піримідину пурину. Інший клас нуклеотидів поміщається в ДНК, і спіраль буде спотворена (особливо з парою основи пурин-пурину). Прикладами є A, що замінює T або C, або C, що замінює A або G.
За еволюційний час швидкість накопичення переходів перевищує швидкість накопичення перетворень.
Вплив мутацій на мРНК
- Missense мутації викликають заміну амінокислоти. Залежно від конкретної заміни, вона може мати, а може і не мати виявлений фенотипічний наслідок. Деякі заміни, наприклад валін для лейцину в положенні, важливому для підтримки a- спіралі, можуть не викликати помітних змін у структурі або функції білка. Інші заміни, такі як валін для глутамату в місці, який змушує гемоглобін полімеризуватися в дезоксигенізованому стані, викликають значну патологію (серповидноклітинна анемія в цьому прикладі).
- Нісенітниці мутації викликають передчасне припинення перекладу. Вони виникають, коли заміщення, введення або видалення генерує стоп-кодон у мРНК у межах області, яка кодує поліпептид у мРНК дикого типу. Вони майже завжди мають серйозні фенотипічні наслідки.
- Мутації зсуву кадрів - це вставки або видалення, які змінюють кадр зчитування мРНК. Вони майже завжди мають серйозні фенотипічні наслідки.
Не всі базові заміни змінюють закодовані амінокислоти
- Заміна бази може призвести до зміни закодованої поліпептидної послідовності, в цьому випадку заміщення називається несинонімічним або немовчазним.
- Якщо заміщення бази відбувається в виродженому місці в кодоні, щоб закодована амінокислота не змінювалася, її називають синонімічної або безшумною заміною.
Приклад:
A C U ‑> A A U є несинонімічною заміною, яка призводить до Thr → Asn
в той час як,
AC U ‑> AC C є синонімічною заміною, яка призводить до Thr → Thr
- Дослідження закономірностей виродження в генетичному коді показує, що несинонімічні заміни відбуваються здебільшого в першій і другій положеннях кодону, тоді як синонімічні заміни відбуваються здебільшого в третій позиції. Однак з цього правила є кілька винятків.
- Загалом, швидкість фіксації синонімічних заміщень у популяції значно більша, ніж швидкість фіксації несинонімічних замін. Це один з найсильніших аргументів на користь моделі нейтральної еволюції, або еволюційного дрейфу, як основної причини замін, що спостерігаються в природних популяціях.