10: Транскрипція: РНК-полімерази
- Page ID
- 7930

Нагадаємо Центральну догму молекулярної біології: ДНК транскрибується в РНК, яка перекладається в білок. Ми будемо висвітлювати матеріал у такому порядку, оскільки це напрямок потоку інформації.
Вступ
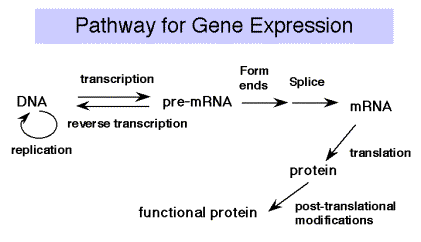
Вивільнений пірофосфат розщеплюється в клітині до 2 Pi, енергетично сприятлива реакція, яка рухає реакцію в напрямку синтезу. При наявності надлишку PPI може відбуватися зворотна реакція пірофосфорилізу. Синтез завжди протікає в напрямку від 5 до 3' (щодо зростаючого ланцюга РНК). Шаблон читається в напрямку від 3 до 5'.
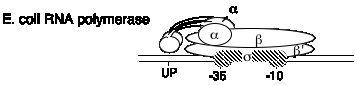
Б. кишкова паличка РНК-полімеразна структура
1. Ця РНК-полімераза синтезує всі класи РНК
мРНК, рРНК, рНК
2. Він складається з чотирьох субодиниць.
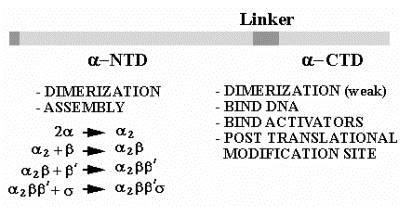
а. ядро і голоензим
А2бб 'а2бб' s
Голофермент = a2bb's = ядро + s = може ініціювати транскрипцію точно, як належний сайт, як визначено промоутером
Ядро = a2bb' = може подовжити зростаючий ланцюг РНК
Промоутера можна визначити двома способами.
- Послідовність ДНК, необхідна для точної, специфічної інітації транскрипції
- Послідовність ДНК, з якою РНК-полімераза зв'язується, щоб точно ініціювати транскрипцію.
б. субодиниці
| Субодиниця | Розмір | Ген | Функція |
|---|---|---|---|
| б' | 160 кДа | РПЦ | b' + b утворюють каталітичний центр. |
| б | 15 кДа | Роб | b' + b утворюють каталітичний центр. |
| a | 40 кДа | Роа | ферментна збірка; також зв'язує послідовність UP в промоторі |
| s | 70 кДа (загальна) | Стрижень | надає специфіку промоутеру; прив'язує до -10 і -35 сайтів в промоутері |
C. кишкова паличка РНК-полімеразний механізм
Спосіб дії s факторів
Наявність s фактора змушує голофермент РНК-полімерази бути селективним у виборі місця ініціації. Це досягається насамперед за рахунок впливу на швидкість дисоціації РНК-полімерази з ДНК.
- а. ядро має сильну спорідненість до загальних послідовностей ДНК. t 1/2 для дисоціації комплексу Core‑ДНК становить близько 60 хв. Це корисно під час фази подовження, але не під час ініціації.
- б. голоензим має знижену спорідненість до загальної ДНК; вона зменшується приблизно в 10 4 рази. t 1/2 для дисоціації голоферменту із загальної ДНК знижується приблизно до 1 сек.
- c. голоензим має значно підвищену спорідненість до послідовностей промоторів. t 1/2 для дисоціації голоферменту з послідовностей промоторів становить близько годин.
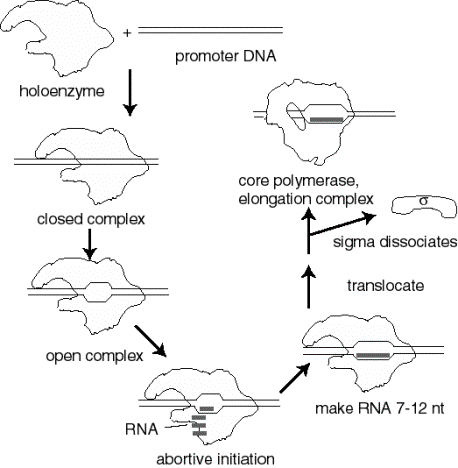
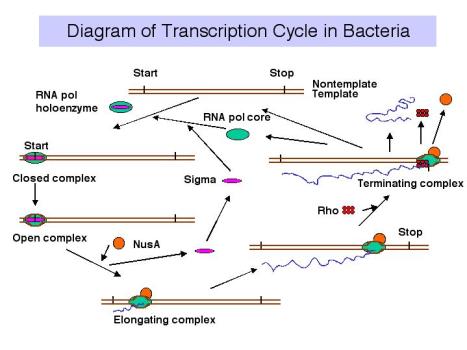
Події при ініціації транскрипції
- а. голофермент РНК-полімерази зв'язується з промотором з утворенням замкнутого комплексу; на цьому етапі не відбувається розмотування ДНК.
- б. полімеразно-промоторний комплекс проходить закритий до відкритого переходу, який являє собою плавлення або розмотування близько 12 б.п.
- c Ініціюючі нуклеотиди можуть зв'язуватися з ферментом, як вказано їх комплементарними нуклеотидами в ланцюжку шаблону ДНК, і фермент буде каталізувати утворення фосфодіефірного зв'язку між ними. Цей комплекс полімерази — ДНК-РНК називають трійчастим комплексом.
- d Під час абортивної ініціації полімераза каталізує синтез коротких стенограм близько 6 або близько того нуклеотидів довжиною, а потім вивільняє їх.
- e Ця фаза закінчується, коли зароджується РНК ~ 6 нуклеотидів зв'язується з другим місцем зв'язування РНК на ферменті; цей другий сайт відрізняється від каталітичного центру. Це зв'язування пов'язане з «скиданням» каталітичного центру так, що фермент тепер буде каталізувати синтез олігонуклеотидів довжиною 7-12.
- f. фермент тепер переходить на нову позицію на шаблоні. Під час цього процесу сигма виходить з комплексу. Конформаційна зміна ферменту, пов'язаного з сигмою, що виходить з комплексу, дозволяє «великий палець» обернути навколо шаблону ДНК, фіксуючи технологічність. Таким чином, основний фермент каталізує синтез РНК під час подовження, яке триває до тих пір, поки не зустрічаються «сигнали», які вказують на припинення.
Малюнок 3.1.8. Події при ініціації
3. Цикл транскрипції
4. Сайти на РНК-полімеразному ядрі
а. фермент охоплює близько 60 б.п. ДНК, при цьому транскрипційний міхур близько 17 bp розмотується.
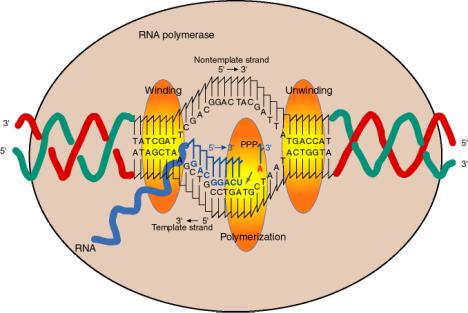
d Вхідний нуклеотид (NTP), який буде доданий до зростаючого ланцюга РНК, зв'язується поруч з 3' кінцем зростаючого ланцюга РНК, за вказівкою шаблону, на активній ділянці для полімеризації.
е Вхідний нуклеотид пов'язаний зі зростаючим ланцюгом РНК нуклеофільної атакою 3' ОН на фосфорил NTP, з вивільненням пірофосфату.
f Реакція прогресує (фермент рухається) близько 50 нц в сек. Це набагато повільніше, ніж швидкість реплікації (близько 1000 нц в сек).
г Якщо шаблон топологічно обмежений, ДНК попереду РНК-полімерази стає перемотаною (позитивні суперспіральні повороти), а ДНК за РНК-полімеразою стає підмотаною (негативні суперспіральні витки).
Ефект розмотування шаблону ДНК РНК-полімеразою полягає в зменшенні T на 1 за кожні 10 bp розмотаного. При цьому DT = -1, а оскільки ДЛ = 0, то DW = +1 за кожні 10 б.п. розмотується. Цей ефект збільшення W буде надаватися в ДНК попереду полімерази.
Ефект перемотування шаблону ДНК РНК-полімеразою, звичайно, навпаки. Т збільшиться на 1 за кожні 10 б.п. перемотування. При цьому ДТ= +1, а оскільки ДЛ = 0, то DW = -1 за кожні 10 б.п. перемотувати. Цей ефект зменшення W буде проявлятися в ДНК за полімеразою, оскільки саме там відбувається перемотування.
5. Інгібітори: корисні реагенти та підказки для функціонування
а. рифаміцин, наприклад рифампіцин: зв'язують субодиницю b, щоб блокувати ініціацію. Препарат запобігає додаванню 3-го або 4-го нуклеотиду, отже, процес ініціації не може бути завершений.
Як ми знаємо, місцем дії рифампіцину є субодиниця b? Мутації, що надають резистентність до рифампіцину, карті гену RPoB.
б. стрептолідигіни: зв'язуються з субодиницею b, щоб пригнічувати подовження ланцюга.
Ці ефекти рифаміцинов і стрептолідигінів, а також те, що вони діють на b субодиницю, стверджують, що субодиниця b потрібна для приєднання нуклеотидів до зростаючого ланцюга.
c Гепарин, поліаніон, зв'язується з b' субодиницею, щоб запобігти зв'язуванню з ДНК in vitro
D. еукаріотичні РНК-полімерази
1. Еукаріоти мають в своїх ядрах 3 різні РНК-полімерази.
- а Кожна ядерна РНК-полімераза являє собою великий білок з приблизно 8-14 субодиницями. МВт становить приблизно 500 000 для кожного.
- б Кожна полімераза має різну функцію:
| РНК-полімераза | локалізація | синтезує | ефект аманітину |
|---|---|---|---|
| РНК-полімераза I | ядерце | Пре-РРНК | жоден |
| РНК-полімераза II | нуклеоплазма | Перед‑МРНК деякі SNRNA | інгібується низькими концентраціями (0,03 мг/мл) |
| РНК-полімераза III | нуклеоплазма | Пре-тРНК, інші малі РНК деякі SNRNA | інгібується високими концентраціями (100 мг/мл) |
2. Структури субодиниць
а Виділено гени та закодовані білки для субодиниць РНК-полімераз дріжджів та визначено послідовності, проведено певний функціональний аналіз.
b Деякі субодиниці гомологічні бактеріальним РНК-полімеразам: Найбільшими двома субодиницями є гомологи b і b'. Приблизно 40 кДа субодиниця є гомологом a.
c Деякі субодиниці є загальними для всіх трьох РНК-полімераз.
d Приклад дріжджової РНК-полімерази II:
| Приблизний розмір (кДа) | субодиниці на полімеразу | роль/коментар | |
|---|---|---|---|
| 220 | 1 | пов'язані з b' | каталітичний? |
| 130 | 1 | пов'язані з б | каталітичний? |
| 40 | 2 | пов'язані з | збірка? |
| 35 | < 1 | ||
| 30 | 2 | загальні для всіх 3 | |
| 27 | 1 | загальні для всіх 3 | |
| 24 | < 1 | ||
| 20 | 1 | загальні для всіх 3 | |
| 14 | 2 | ||
| 10 | 1 |
e. Найбільша субодиниця має карбокситермінальну область (КТД) з незвичайною структурою: тандемні повторення послідовності Тир-Сер‑Про-Thr‑Ser‑Pro‑Ser‑Pro‑Thr. Дріжджовий фермент має 26 тандемних повторів, а фермент ссавців - близько 50. Вони можуть бути фофорильовані на Ser і Thr, щоб дати сильно заряджений CTD.
- РНК Пол IIa не фосфорилюється в КТД.
- РНК Pol IiO фосфорилюється в КТД.

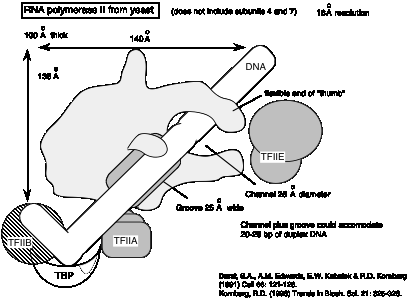
4. РНК-полімерази в хлоропластах (пластидах) і мітохондріях
- а РНК-полімераза, виявлена в пластидах, кодується на пластидній хромосомі. У деяких видів мітохондріальна РНК-полімераза кодується мітохондріальної ДНК.
- б Ці органеллярні РНК-полімерази набагато більше пов'язані з бактеріальними РНК-полімеразами, ніж з ядерними РНК-полімеразами. Це є вагомим аргументом на користь того, що ці органели є бактеріальними, підтримуючи ендо-симбіотну модель для придбання цих органел у еукаріотів.
- c Ці РНК-полімерази каталізують специфічну транскрипцію органелярних генів.
Загальні фактори транскрипції еукаріотичної РНК-полімерази II
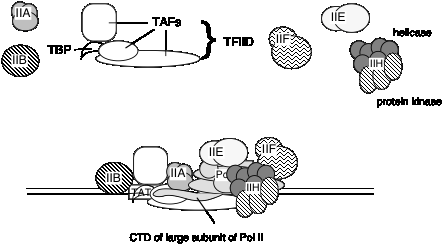
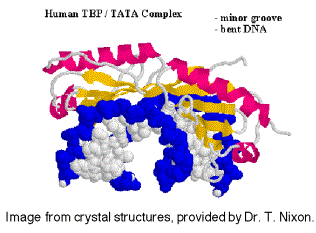
Невідомо, чи є один і той же набір TAF в TFIID для всіх промоутерів, транскрибуваних РНК-полімеразою II, або якщо деякі використовуються лише для певних типів промоутерів. TFIID є єдиним специфічним для послідовності фактором загальної транскрипції, який досі характеризується, і він зв'язується у незначній канавці ДНК. Він також використовується в промоутерах Tata less, тому роль прив'язки до конкретної послідовності все ще знаходиться під дослідженням.
3. Короткий зміст загальних факторів транскрипції для РНК-полімерази II.
Фактори РНК-полімерази II (клітини людини)
| Фактор | Кількість субодиниць | Молекулярний маса (кДа) | Функції | Функції для Рекрут: |
|---|---|---|---|---|
| ШИРИНА: СТ.Л. | 1 | 38 | Розпізнати основний промоутер (TATA) | ТІБІБ |
| TID: Теґи | 12 | 15-250 | Розпізнати основний промоутер (Non-TATA); Позитивне та негативне регулювання | РНК Пол II? |
| TFIIA | 2 | 12, 19, 35 | Стабілізувати зв'язування TBP-ДНК; Анти-репресії | |
| ТІБІБ | 1 | 35 | Виберіть стартовий сайт для РНК Pol II | РНК Поліі-TFIIF |
| РНК Пол II | 12 | 10-220 | Каталізувати синтез РНК | ТРИДЦЯТЬ |
| TFIIF | 2 | 30, 74 | Цільова РНК POII на промотор; дестабілізація неспецифічних взаємодій між POII та ДНК | |
| ТРИДЦЯТЬ | 2 | 34, 57 | Модулювати TFIIH хелікази, АТФази та кінази активності; Безпосередньо посилити промоторне плавлення? | TFIIH |
| TFIIH | 9 | 35-89 | Helicase розплавити промотор; CTD кіназа; промоутер кліренс? |
Редер, Р.Г. (1996) ТІБС 21:327-335.
4. TFIIH - це мультисубодиничний фактор транскрипції, який також бере участь у репарації ДНК.
Субодиниці людського фактора
| Ген | Молець. маса білка (кДа) | Функція/Структура | Запропонована роль |
|---|---|---|---|
| XPB | 89 | helicase, треки 3' до 5' | Розмотувати дуплекс для транскрипції/ремонту |
| XPD | 80 | helicase, треки 5' до 3' | Розмотувати дуплекс, Ремонт |
| P62 | 62 | невідомий | |
| Р52 | 52 | невідомий | |
| P44 | 44 | Zn-палець | Зв'язує ДНК |
| Р34 | 34 | Zn-палець | |
| МАТ 1 | 32 | CDK коефіцієнт збірки | |
| Циклін Н | 38 | Велосипедний партнер для CDK7/MO15 | |
| ЦДК7/МО15 | 32 | протеїнкіназа | Кіназа для КТД |

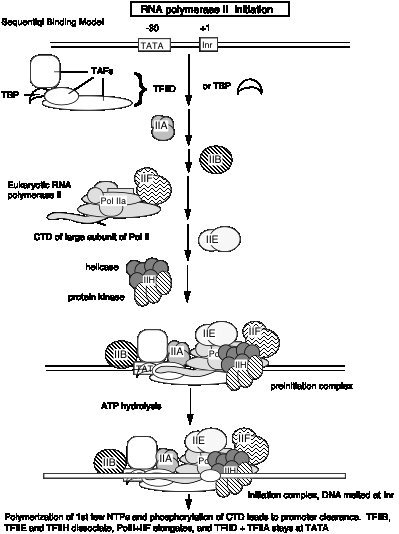
Таблиця 3.1.6. РНК-полімераза II голофермент і медіатор

Ці дослідження показують, що РНК-полімераза II може існувати в декількох різних станах або комплексах. Один знаходиться в дуже великому голокомплексі, що містить медіатор. У такому стані він точно ініціює транскрипцію при призначенні TFIID, і реагує на активатори (табл. 3.1.6). Медіаторний субкомплекс, як видається, здатний дисоціювати та реасоціюватися з РНК-полімеразою II та ГТФ. Дійсно, ця реасоціація може бути кроком, який був застосований у ідентифікації посередника. Без медіатора РНК-полімераза II плюс GTF можуть ініціювати транскрипцію в правильному місці (за вказівкою TFIID), але вони не реагують на активатори. При відсутності ГТФ РНК-полімераза II здатна транскрибувати шаблони ДНК, але вона не почне транскрипцію на правильному місці. Отже, він компетентний для подовження, але не ініціації.
Таблиця 3.1.7. Розширення функцій РНК-полімерази II
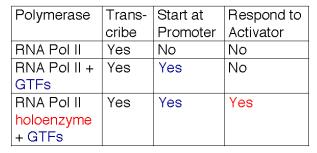
Рис.3.1.17
![]()
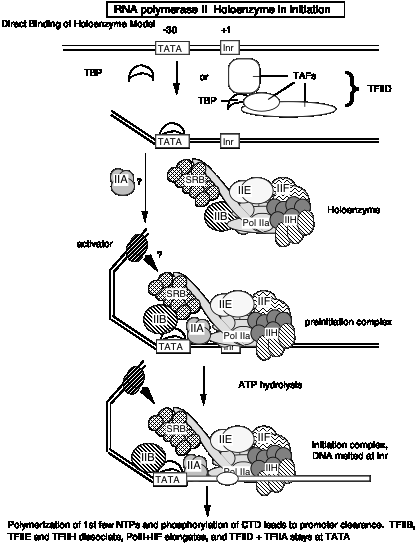
Якщо голофермент є первинним ферментом, який бере участь у ініціації транскрипції в еукаріотичних клітині, то прогресивний шлях складання, що спостерігається in vitro (див. Розділ d вище), може мати незначне значення in vivo. Можливо, холоензим зв'яжеться з промоторами, просто позначені зв'язуванням TBP (або TFIID) з коробкою TATA, на відміну від прогресивної моделі складання, яка має більш широкий, упорядкований механізм складання. В обох моделах прив'язка TBP або TFIID є початковим етапом складання комплексу преініціації. Однак на даний момент не можна виключати можливості того, що холоензим використовується в одних промоторах, а прогресивна збірка відбувається в інших.
7. Мішені для білків-активаторів
Мішенями для білків-активаторів транскрипції може бути якийсь компонент комплексу ініціації. Одним з напрямків розслідування є вказівка на TAF в TFIID, а також на TFIIB як цілі для активаторів. Таким чином, активатори можуть полегшити впорядковану збірку комплексу ініціації шляхом набору GTF. Однак холоензим містить «медіатор» або комплекс SRB, який може опосередковувати реакцію на активатори. Таким чином, активатори можуть служити для набору голоферменту в промотор. Потрібні подальші дослідження, щоб встановити, чи правильний той чи інший, чи це окремі шляхи до активації.
F. загальні фактори транскрипції еукаріотичних РНК-полімераз I та III
1. Загальні фактори транскрипції РНК-полімерази I
а. основний промоутер охоплює стартовий сайт транскрипції, плюс елемент управління вище за течією розташований приблизно 70 bp далі 5'.
b Фактор UBF1 зв'язується з багатою послідовністю G+C як в елементі управління вище за течією, так і в основному промоутері.
c Мультисубодиничний комплекс, який називається SL1, зв'язується з комплексом UBF1‑ДНК, знову як на вищому, так і на основному елементах.
d Однією з субодиниць SL1 є TBP ‑ білок, що зв'язує TATA з TFIID!
е РНК-полімераза I потім зв'язується з цим комплексом ДНК+UBF1+SL1, щоб ініціювати транскрипцію при правильному нуклеотиді і подовженому, щоб зробити pre‑RRNA.
2. Загальні фактори транскрипції для РНК Pol III
- а Внутрішні контрольні послідовності характерні для генів, транскрибуваних РНК Pol III (див. нижче).
- b. tfIIIA: зв'язується з областю внутрішнього контролю генів, які кодують 5S РНК (внутрішній промотор типу 1)
- c. TFIIIIC: зв'язується з областями внутрішнього контролю генів для 5S РНК (поряд з TFIIIIA) та для ТРНК (внутрішні промотори типу 2)
- d. TFIIIB: Прив'язка TFIIIIC спрямовує TFIIIB на прив'язку до послідовностей (від -40 до +11), які перекривають початкову ділянку для транскрипції. Однією субодиницею TFIIIB є TBP, хоча для транскрипції не потрібно поле TATA. TFIIIIA та TFIIIIC тепер можуть бути видалені, не впливаючи на здатність РНК-полімерази III ініціювати транскрипцію. Таким чином, ТФІІІА та ТФІІІК є факторами складання, а ТФІІІБ — фактором ініціації.
Малюнок 3.1.18.
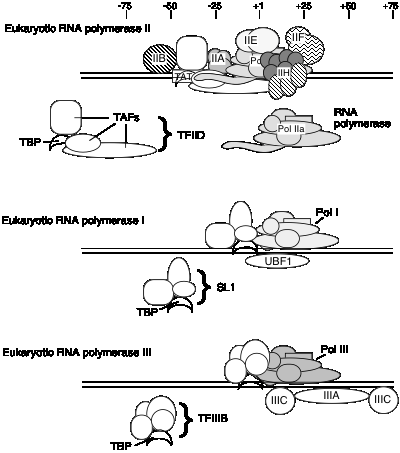
е. РНК-полімераза III зв'язується з комплексом TFIIIB+ДНК для точної та ефективної транскрипції.
3 Фактор транскрипції, який використовується всіма 3 РНК полязами: ТБП
ТБП, здається, відіграє спільну роль у напрямку РНК-полімерази (I, II та III) для ініціювання у правильному місці. Багатосубодиничні фактори, що містять TBP (TFIID, SL1 та TFIIIB), можуть служити факторами позиціонування відповідних полімераз.