9.1: Сигнальні молекули та клітинні рецептори
- Page ID
- 1812

Навички для розвитку
- Опишіть чотири типи сигналізації, виявлені в багатоклітинних організмах
- Порівняйте внутрішні рецептори з рецепторами поверхні клітин
- Визнати взаємозв'язок між будовою ліганда та механізмом його дії
У світі живих клітин існує два види зв'язку. Зв'язок між клітинами називається міжклітинної сигналізацією, а зв'язок всередині клітини називається внутрішньоклітинної сигналізацією. Простий спосіб запам'ятати відмінність - зрозуміти латинське походження приставок: inter- означає «між» (наприклад, перетинаються лінії - це ті, що перетинаються один одного), а intra- означає «всередині» (як внутрішньовенно).
Хімічні сигнали виділяються сигнальними клітинами у вигляді невеликих, зазвичай летючих або розчинних молекул, званих лігандами. Ліганд - це молекула, яка зв'язує іншу специфічну молекулу, в деяких випадках доставляючи сигнал в процесі. Таким чином, ліганди можна розглядати як сигнальні молекули. Ліганди взаємодіють з білками в клітинах-мішенях, які є клітинами, на які впливають хімічні сигнали; ці білки також називають рецепторами. Ліганди та рецептори існують у декількох різновидах; однак конкретний ліганд матиме специфічний рецептор, який зазвичай зв'язує лише цей ліганд.
Форми сигналізації
Існує чотири категорії хімічної сигналізації, виявленої в багатоклітинних організмах: паракринна сигналізація, ендокринна сигналізація, аутокринна сигналізація та пряма сигналізація через розриви переходів (рис.\(\PageIndex{1}\)). Основна відмінність між різними категоріями сигналізації полягає в відстані, яке сигнал проходить через організм, щоб досягти клітини-мішені. Не на всі клітини впливають одні й ті ж сигнали.
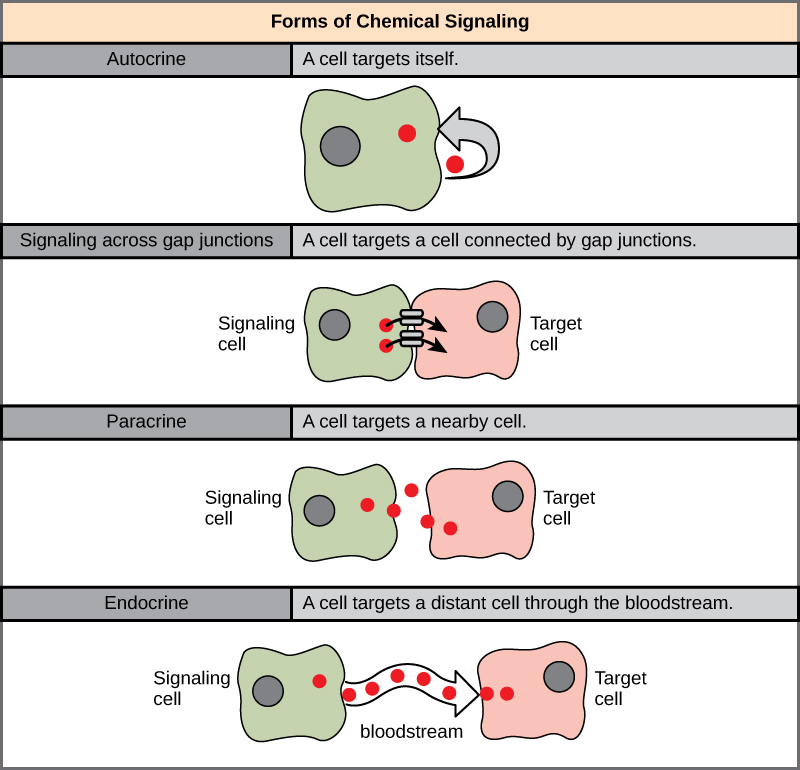
Паракринна сигналізація
Сигнали, які діють локально між клітинами, які знаходяться близько один до одного, називаються паракринними сигналами. Паракринні сигнали рухаються шляхом дифузії через позаклітинний матрикс. Ці типи сигналів зазвичай викликають швидкі відповіді, які тривають лише короткий проміжок часу. Для того, щоб зберегти відповідь локалізованою, молекули паракринного ліганду зазвичай швидко деградуються ферментами або видаляються сусідніми клітинами. Видалення сигналів відновить градієнт концентрації для сигналу, що дозволить їм швидко розсіюватися через внутрішньоклітинний простір, якщо вони знову звільняються.
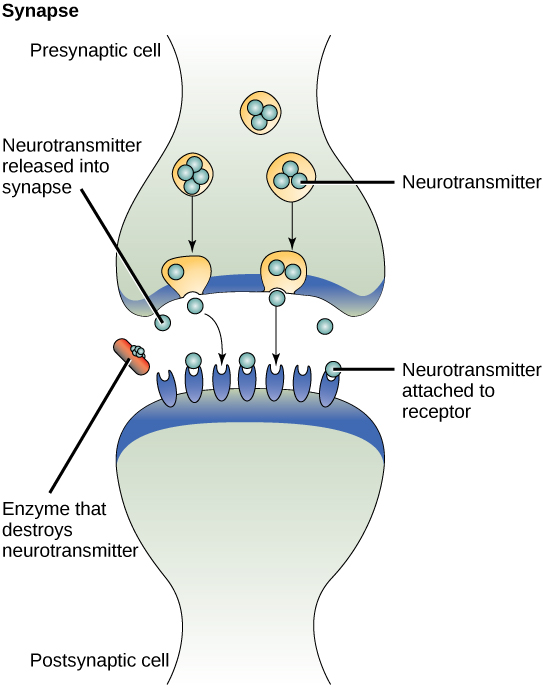
Одним із прикладів паракринної сигналізації є передача сигналів через синапси між нервовими клітинами. Нервова клітина складається з клітинного тіла, декількох коротких, розгалужених розширень, які називаються дендритами, які отримують подразники, і довгого розширення, яке називається аксоном, який передає сигнали іншим нервовим клітинам або м'язовим клітинам. Місце з'єднання між нервовими клітинами, де відбувається передача сигналу, називається синапсом. Синаптичний сигнал - це хімічний сигнал, який рухається між нервовими клітинами. Сигнали всередині нервових клітин поширюються швидкоплинними електричними імпульсами. Коли ці імпульси досягають кінця аксона, сигнал продовжується до дендриту наступної клітини шляхом вивільнення хімічних лігандів, званих нейромедіаторами пресинаптичної клітиною (клітиною, що випромінює сигнал). Нейромедіатори транспортуються на дуже малі відстані між нервовими клітинами, які називаються хімічними синапсами (рис.\(\PageIndex{2}\)). Невелика відстань між нервовими клітинами дозволяє сигналу швидко подорожувати; це дозволяє негайно реагувати, наприклад, Візьміть руку з плити!
Коли нейромедіатор зв'язує рецептор на поверхні постсинаптичної клітини, електрохімічний потенціал клітини-мішені змінюється, і запускається наступний електричний імпульс. Нейромедіатори, які виділяються в хімічний синапс, швидко деградують або реабсорбуються пресинаптичною клітиною, щоб нервова клітина реципієнта могла швидко відновитися і бути готовою швидко реагувати на наступний синаптичний сигнал.
Ендокринна сигналізація
Сигнали віддалених клітин називаються ендокринними сигналами, і вони походять з ендокринних клітин. (В організмі багато ендокринних клітин розташовані в ендокринних залозах, таких як щитовидна залоза, гіпоталамус і гіпофіз.) Ці типи сигналів зазвичай виробляють більш повільну реакцію, але мають більш тривалий ефект. Ліганди, що випускаються в ендокринній сигналізації, називаються гормонами, сигналізуючи молекули, які виробляються в одній частині тіла, але впливають на інші області тіла на деякій відстані.
Гормони подорожують великі відстані між ендокринними клітинами та їх клітинами-мішенями через кров, що є відносно повільним способом переміщення по всьому тілу. Через форму транспорту гормони розбавляються і присутні в низьких концентраціях, коли вони діють на клітини-мішені. Цим відрізняється від паракринної сигналізації, при якій місцеві концентрації лігандів можуть бути дуже високими.
Автокринна сигналізація
Аутокринні сигнали виробляються сигнальними клітинами, які також можуть зв'язуватися з вивільняється лігандом. Це означає, що сигнальна комірка і цільова клітина можуть бути однаковими або схожими осередками (префікс авто- означає себе, нагадування про те, що сигнальна комірка посилає сигнал собі). Цей тип сигналізації часто виникає під час раннього розвитку організму, щоб забезпечити розвиток клітин в правильні тканини і взяти на себе належну функцію. Аутокринна сигналізація також регулює больові відчуття і запальні реакції. Далі, якщо клітина заражена вірусом, клітина може сигналізувати собі про запрограмовану загибель клітин, вбиваючи вірус в процесі. У деяких випадках на сусідні клітини того ж типу впливає і звільнився ліганд. При ембріологічному розвитку цей процес стимуляції групи сусідніх клітин може допомогти спрямувати диференціювання однакових клітин в один і той же тип клітин, забезпечуючи тим самим належний результат розвитку.
Пряма сигналізація через зазорні переходи
Розривні переходи у тварин і плазмодезмати у рослин є зв'язками між плазматичними мембранами сусідніх клітин. Ці наповнені водою канали дозволяють малим сигнальним молекулам, званим внутрішньоклітинними медіаторами, дифузно між двома клітинами. Дрібні молекули, такі як іони кальцію (Ca 2+), здатні переміщатися між клітинами, але великі молекули, такі як білки та ДНК, не можуть вміститися через канали. Специфіка каналів гарантує, що клітини залишаються незалежними, але можуть швидко і легко передавати сигнали. Передача сигнальних молекул повідомляє про поточний стан клітини, яка знаходиться безпосередньо поруч з клітиною-мішенню; це дозволяє групі клітин координувати свою реакцію на сигнал, який міг отримати тільки один з них. У рослин плазмодесмати поширені повсюдно, перетворюючи всю рослину в гігантську, комунікаційну мережу.
типи рецепторів
Рецептори - це молекули білка в клітині-мішені або на її поверхні, які зв'язують ліганд. Існує два типи рецепторів, внутрішні рецептори і рецептори клітинної поверхні.
внутрішні рецептори
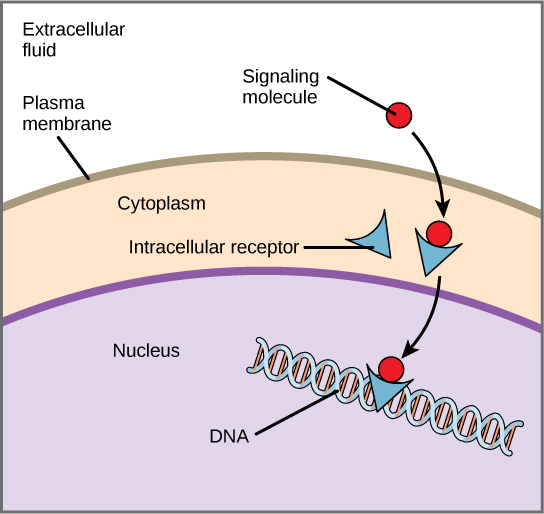
Внутрішні рецептори, також відомі як внутрішньоклітинні або цитоплазматичні рецептори, знаходяться в цитоплазмі клітини і реагують на гідрофобні молекули ліганду, які здатні подорожувати через плазматичну мембрану. Потрапивши всередину клітини, багато з цих молекул зв'язуються з білками, які діють як регулятори синтезу (транскрипції) мРНК для опосередковування експресії генів. Експресія генів - це клітинний процес перетворення інформації в ДНК клітини в послідовність амінокислот, яка в кінцевому підсумку утворює білок. Коли ліганд зв'язується з внутрішнім рецептором, спрацьовує конформаційна зміна, що викриває на білку ділянку зв'язування ДНК. Комплекс ліганд-рецепторів переміщається в ядро, потім зв'язується з конкретними регуляторними областями хромосомної ДНК і сприяє ініціації транскрипції (рис.\(\PageIndex{3}\)). Транскрипція - це процес копіювання інформації в ДНК клітини в спеціальну форму РНК, звану месенджерною РНК (мРНК); клітина використовує інформацію в мРНК (яка рухається в цитоплазму і асоціюється з рибосомами), щоб зв'язати конкретні амінокислоти в правильному порядку, виробляючи білок. Внутрішні рецептори можуть безпосередньо впливати на експресію генів без необхідності передавати сигнал іншим рецепторам або месенджерам.
Поверхневі рецептори клітин
Поверхневі рецептори клітин, також відомі як трансмембранні рецептори, - це клітинна поверхня, мембранно-закріплені (інтегральні) білки, які зв'язуються з зовнішніми молекулами ліганду. Цей тип рецепторів охоплює плазматичну мембрану і здійснює передачу сигналу, при якій позаклітинний сигнал перетворюється в міжклітинний сигнал. Ліганди, які взаємодіють з рецепторами поверхні клітин, не повинні потрапляти в клітину, на яку вони впливають. Поверхневі рецептори клітин також називають клітинно-специфічними білками або маркерами, оскільки вони специфічні для окремих типів клітин.
Оскільки білки рецепторів поверхні клітин є основоположними для нормального функціонування клітин, не дивно, що збій у будь-якому з цих білків може мати серйозні наслідки. Показано, що помилки в білкових структурах певних молекул рецепторів відіграють певну роль при гіпертонії (високий кров'яний тиск), астмі, захворюваннях серця та раку.
Кожен рецептор поверхні клітини має три основні компоненти: зовнішній ліганд-зв'язуючий домен, гідрофобну мембрану, що охоплює область, і внутрішньоклітинний домен всередині клітини. Ліганд-зв'язуючий домен також називають позаклітинним доменом. Розмір і протяжність кожного з цих доменів варіюються в широких межах, залежно від типу рецептора.
Еволюція з'єднання: як віруси розпізнають хоста
На відміну від живих клітин, багато вірусів не мають плазматичної мембрани або будь-якої з структур, необхідних для підтримки життя. Деякі віруси просто складаються з інертної білкової оболонки, що містить ДНК або РНК. Для розмноження віруси повинні вторгнутися в живу клітину, яка служить господарем, а потім взяти на себе господарів клітинний апарат. Але як вірус розпізнає свого господаря?
Віруси часто зв'язуються з рецепторами поверхні клітин на клітині-господаря. Наприклад, вірус, що викликає грип людини (грип), зв'язується саме з рецепторами на мембранах клітин дихальної системи. Хімічні відмінності рецепторів поверхні клітин у господарів означають, що вірус, який заражає конкретний вид (наприклад, людину), не може заразити інший вид (наприклад, курей).
Однак віруси мають дуже малу кількість ДНК або РНК порівняно з людиною, і, як наслідок, розмноження вірусів може відбуватися швидко. Розмноження вірусів незмінно спричиняє помилки, які можуть призвести до змін у знову вироблених вірусах; ці зміни означають, що вірусні білки, які взаємодіють з рецепторами поверхні клітин, можуть розвиватися таким чином, що вони можуть зв'язуватися з рецепторами у нового господаря. Такі зміни відбуваються випадковим чином і досить часто в репродуктивному циклі вірусу, але зміни мають значення тільки в тому випадку, якщо вірус з новими зв'язуючими властивостями вступає в контакт з відповідним господарем. У разі грипу така ситуація може виникнути в умовах, де тварини та люди знаходяться в тісному контакті, таких як птахівництво та свиноферми. 1 Як тільки вірус перестрибує на нового господаря, він може швидко поширитися. Вчені уважно спостерігають за новоявленими вірусами (званими з'являються вірусами) в надії, що такий моніторинг може знизити ймовірність глобальних вірусних епідемій.
Клітинно-поверхневі рецептори беруть участь у більшій частині сигналізації в багатоклітинних організмах. Існує три загальні категорії рецепторів поверхні клітин: рецептори, пов'язані з іонними каналами, рецептори, пов'язані з G-білками, і рецептори, пов'язані з ферментами.
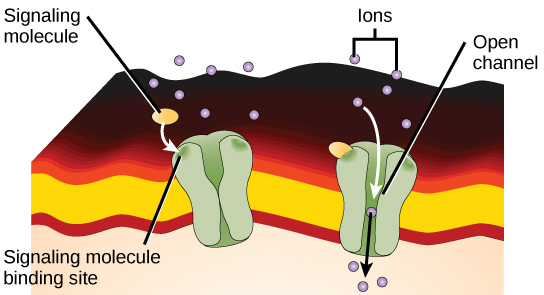
Рецептори, пов'язані з іонним каналом, пов'язують ліганд і відкривають канал через мембрану, що дозволяє проходити специфічні іони. Для формування каналу цей тип рецептора поверхні клітин має велику область, що охоплює мембрану. Для того, щоб взаємодіяти з хвостами фосфоліпідних жирних кислот, які утворюють центр плазматичної мембрани, багато амінокислот в області мембрани мають гідрофобну природу. І навпаки, амінокислоти, які вистилають внутрішню частину каналу, є гідрофільними, щоб забезпечити проходження води або іонів. Коли ліганд зв'язується з позаклітинною областю каналу, відбувається конформаційна зміна структури білків, що дозволяє проходити через них іони, такі як натрій, кальцій, магній і водень (рис.\(\PageIndex{4}\)).
G-білкові рецептори зв'язують ліганд і активують мембранний білок, який називається G-білком. Активований G-білок потім взаємодіє або з іонним каналом, або з ферментом в мембрані (рис.\(\PageIndex{5}\)). Всі рецептори, пов'язані з G-білками, мають сім трансмембранних доменів, але кожен рецептор має свій специфічний позаклітинний домен і сайт, що зв'язує G-білок.
Клітинна сигналізація за допомогою G-білкових рецепторів відбувається у вигляді циклічного ряду подій. Перш ніж ліганд зв'язується, неактивний G-білок може зв'язуватися з нововиявленим ділянкою на рецепторі, специфічному для його зв'язування. Як тільки G-білок зв'язується з рецептором, результуюча зміна форми активує G-білок, який вивільняє ВВП і підхоплює GTP. Субодиниці G-білка потім розщеплюються на α субодиницю і βγ субодиницю. Один або обидва ці фрагменти G-білка можуть в результаті активувати інші білки. Через деякий час ГТП на активній α субодиниці G-білка гідролізується до ВВП і βγ субодиниця деактивується. Субодиниці переасоціюються з утворенням неактивного G-білка і цикл починається заново.
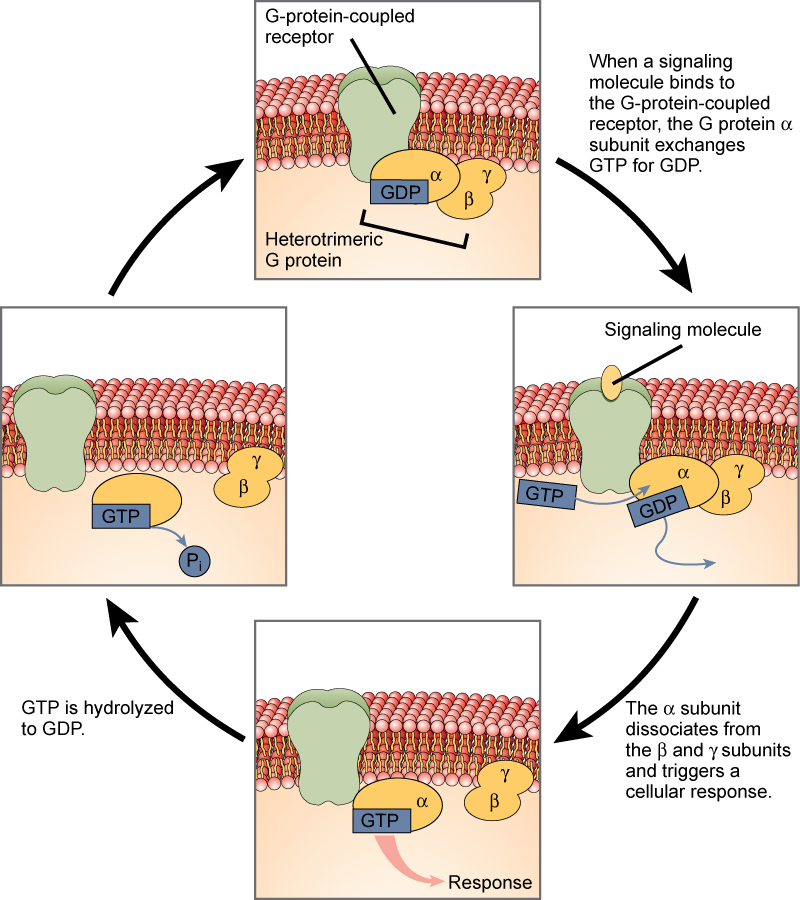
G-білкові рецептори були широко вивчені, і багато було вивчено про їх роль у підтримці здоров'я. Бактерії, які є патогенними для людини, можуть виділяти отрути, які переривають специфічну функцію рецепторів G-білків, що призводить до таких захворювань, як коклюш, ботулізм та холера. Наприклад, при холері (рис.\(\PageIndex{6}\)) Бактерія, що переноситься водою Vibrio cholerae, виробляє токсин, холераген, який зв'язується з клітинами, що вистилають тонку кишку. Потім токсин потрапляє в ці клітини кишечника, де модифікує G-білок, який контролює відкриття хлоридного каналу і змушує його залишатися постійно активним, що призводить до великих втрат рідини з організму і потенційно смертельного зневоднення в результаті.

Ферментно-зв'язані рецептори - це рецептори поверхні клітин з внутрішньоклітинними доменами, які пов'язані з ферментом. У деяких випадках внутрішньоклітинний домен самого рецептора є ферментом. Інші рецептори, пов'язані з ферментами, мають невеликий внутрішньоклітинний домен, який взаємодіє безпосередньо з ферментом. Ферментно-зв'язані рецептори зазвичай мають великі позаклітинні та внутрішньоклітинні домени, але мембранно-охоплююча область складається з однієї альфа-спіральної області пептидного пасма. Коли ліганд зв'язується з позаклітинним доменом, через мембрану передається сигнал, активуючи фермент. Активація ферменту запускає ланцюжок подій всередині клітини, що в кінцевому підсумку призводить до відповіді. Одним із прикладів цього типу рецептора, зв'язаного з ферментами, є рецептор тирозинкінази (рис.\(\PageIndex{7}\)). Кіназа - це фермент, який переносить фосфатні групи з АТФ на інший білок. Рецептор тирозинкінази переносить фосфатні групи в молекули тирозину (залишки тирозину). По-перше, сигнальні молекули зв'язуються з позаклітинним доменом двох прилеглих рецепторів тирозинкінази. Потім два сусідніх рецептора зв'язуються між собою або димеризуються. Потім фосфати додають до залишків тирозину на внутрішньоклітинному домені рецепторів (фосфорилювання). Потім фосфорильовані залишки можуть передавати сигнал наступному месенджеру в цитоплазмі.
Мистецтво З'єднання
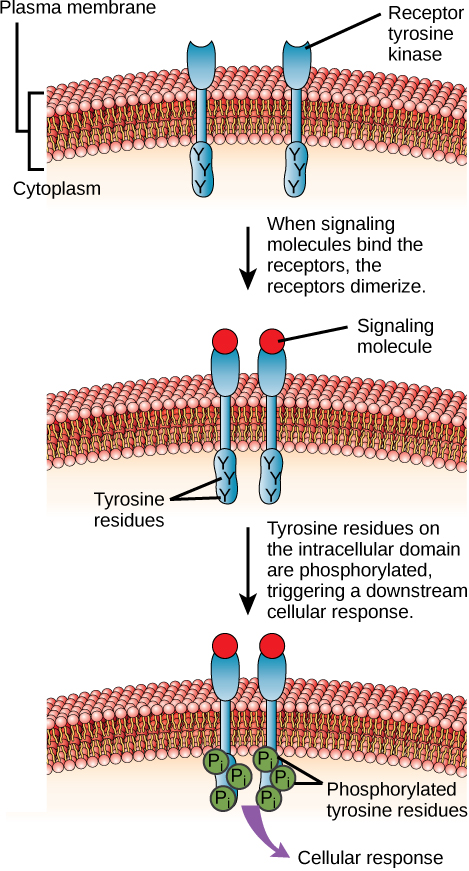
HER2 є рецептором тирозинкінази. У 30 відсотках раку молочної залози людини HER2 постійно активується, що призводить до нерегульованого поділу клітин. Лапатініб, препарат, який використовується для лікування раку молочної залози, пригнічує аутофосфорилювання рецепторів тирозинкінази HER2 (процес, за допомогою якого рецептор додає фосфати на себе), тим самим зменшуючи ріст пухлини на 50 відсотків. Крім аутофосфорилювання, який з наступних кроків буде пригнічений лапатінібом?
- Сигналізація зв'язування молекул, димеризація та клітинна реакція нижче за течією
- Димеризація та клітинна реакція нижче за течією
- Клітинна реакція нижче за течією
- Активність фосфатази, димеризація та клітинна реакція нижньої пари
Сигнальні молекули
Вироблені сигнальними клітинами і подальшим зв'язуванням з рецепторами в клітинах-мішенях, ліганди діють як хімічні сигнали, які рухаються до клітини-мішеней для координації відповідей. Типи молекул, які служать лігандами, неймовірно різноманітні і варіюються від невеликих білків до малих іонів, таких як кальцій (Ca 2+).
Малі гідрофобні ліганди
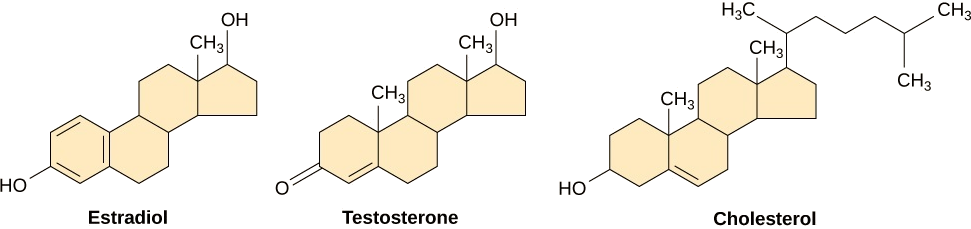
Дрібні гідрофобні ліганди можуть безпосередньо дифузіровать через плазматичну мембрану і взаємодіяти з внутрішніми рецепторами. Важливими членами цього класу лігандів є стероїдні гормони. Стероїди - це ліпіди, які мають вуглеводневий скелет з чотирма плавленими кільцями; різні стероїди мають різні функціональні групи, прикріплені до вуглецевого скелета. Стероїдні гормони включають жіночий статевий гормон, естрадіол, який є різновидом естрогену; чоловічий статевий гормон, тестостерон; і холестерин, який є важливим структурним компонентом біологічних мембран і попередником стероїдних гормонів (рис.\(\PageIndex{8}\)). Інші гідрофобні гормони включають гормони щитовидної залози та вітамін D. Для того, щоб бути розчинними в крові, гідрофобні ліганди повинні зв'язуватися з білками-носіями, поки вони транспортуються через кров.
Водорозчинні ліганди
Водорозчинні ліганди полярні і тому не можуть проходити через плазматичну мембрану без сторонньої допомоги; іноді вони занадто великі, щоб взагалі проходити через мембрану. Натомість більшість водорозчинних лігандів зв'язуються з позаклітинним доменом клітинно-поверхневих рецепторів. Ця група лігандів досить різноманітна і включає в себе дрібні молекули, пептиди, білки.
Інші ліганди
Оксид азоту (NO) - це газ, який також діє як ліганд. Він здатний дифузувати безпосередньо по плазматичній мембрані, і одна з його ролей полягає у взаємодії з рецепторами в гладкій мускулатурі і індукувати розслаблення тканини. NO має дуже короткий період напіврозпаду і тому функціонує лише на коротких відстанях. Нітрогліцерин, лікування серцевих захворювань, діє, запускаючи вивільнення NO, що змушує кровоносні судини розширюватися (розширюватися), тим самим відновлюючи приплив крові до серця. NO став більш відомим останнім часом, оскільки шлях, який він впливає, орієнтований на ліки, що відпускаються за рецептом для еректильної дисфункції, такі як Віагра (ерекція включає розширені кровоносні судини).
Резюме
Клітини спілкуються як за допомогою міжклітинної, так і внутрішньоклітинної сигналізації. Сигнальні клітини виділяють ліганди, які зв'язуються з клітинами-мішенями і ініціюють ланцюжок подій всередині клітини-мішені. Чотири категорії сигналізації в багатоклітинних організмах - паракринна сигналізація, ендокринна сигналізація, аутокринна сигналізація та пряма сигналізація через розриви переходів. Паракринна сигналізація відбувається на невеликих відстанях. Ендокринні сигнали переносяться на великі відстані по кровотоку гормонами, а аутокринні сигнали приймаються тією ж клітиною, яка посилала сигнал або іншими сусідніми клітинами такого ж роду. Зазорні переходи дозволяють малим молекулам, включаючи сигнальні молекули, протікати між сусідніми клітинами.
Внутрішні рецептори знаходяться в цитоплазмі клітини. Тут вони пов'язують молекули лігандів, які перетинають плазматичну мембрану; ці рецепторно-лігандні комплекси рухаються до ядра і взаємодіють безпосередньо з клітинною ДНК. Клітинно-поверхневі рецептори передають сигнал ззовні клітини в цитоплазму. Іонно-зв'язані рецептори, зв'язуючись зі своїми лігандами, утворюють пору через плазматичну мембрану, через яку можуть проходити певні іони. G-білкові рецептори взаємодіють з G-білком на цитоплазматичній стороні плазматичної мембрани, сприяючи обміну зв'язаного ВВП на ГТП і взаємодіючи з іншими ферментами або іонними каналами для передачі сигналу. Ферментно-зв'язані рецептори передають сигнал ззовні клітини до внутрішньоклітинного домену мембранно-зв'язаного ферменту. Зв'язування лігандів викликає активацію ферменту. Дрібні гідрофобні ліганди (як і стероїди) здатні проникати через плазматичну мембрану і зв'язуватися з внутрішніми рецепторами. Водорозчинні гідрофільні ліганди не в змозі пройти через мембрану; натомість вони зв'язуються з рецепторами поверхні клітин, які передають сигнал всередину клітини.
Мистецькі зв'язки
Малюнок\(\PageIndex{7}\): HER2 є рецептором тирозинкінази. У 30 відсотках раку молочної залози людини HER2 постійно активується, що призводить до нерегульованого поділу клітин. Лапатініб, препарат, який використовується для лікування раку молочної залози, пригнічує аутофосфорилювання рецепторів тирозинкінази HER2 (процес, за допомогою якого рецептор додає фосфати на себе), тим самим зменшуючи ріст пухлини на 50 відсотків. Крім аутофосфорилювання, який з наступних кроків буде пригнічений лапатінібом?
- Сигналізація зв'язування молекул, димеризація та клітинна реакція нижче за течією.
- Димеризація та клітинна реакція нижче за течією.
- Клітинна реакція нижче за течією.
- Активність фосфатази, димеризація і спад клітинної реакції.
- Відповідь
-
C. нижча клітинна реакція буде гальмована.
Виноски
- 1 Сігалов А.Б., Школа природи. IV. Навчання вірусам, Само/Нея 1, № 4 (2010): 282-298. Y.Cao, X. Koh, L. Dong, X. du, A. Wu, X. ding, H. deng, Y. shu, J. Chen, T. jiang, швидка оцінка зв'язуючої активності вірусу грипу гемаглютиніну до рецепторів людини і птахів, PLoS One 6, no. 4 (2011): e18664.
Глосарій
- аутокринний сигнал
- сигнал, який надсилається та приймається тими ж або подібними сусідніми клітинами
- клітинний поверхневий рецептор
- білок поверхні клітини, який передає сигнал від зовнішньої частини клітини всередину, навіть якщо ліганд не потрапляє в клітину
- хімічний синапс
- невеликий простір між аксоновими терміналами та дендритами нервових клітин, де функціонують нейромедіатори
- ендокринна клітина
- клітина, яка виділяє ліганди, що беруть участь в ендокринній сигналізації (гормони)
- ендокринний сигнал
- сигнал на великі відстані, який доставляється лігандами (гормонами), що подорожують через кровоносну систему організмів від сигнальної клітини до клітини-мішені
- ферментно-зв'язаний рецептор
- клітинно-поверхневий рецептор з внутрішньоклітинними доменами, які пов'язані з мембрано-зв'язаними ферментами
- позаклітинний домен
- область рецептора клітинної поверхні, яка розташована на поверхні клітини
- G-білково-зв'язаний рецептор
- клітинно-поверхневий рецептор, який активує мембранно-зв'язані G-білки для передачі сигналу від рецептора до сусідніх мембранних компонентів
- міжклітинна сигналізація
- зв'язок між клітинами
- внутрішній рецептор
- (Також внутрішньоклітинний рецептор) рецепторний білок, який знаходиться в цитозолі клітини і зв'язується з лігандами, які проходять через плазматичну мембрану
- внутрішньоклітинний медіатор
- (також, другий месенджер) мала молекула, яка передає сигнали всередині клітини
- внутрішньоклітинна сигналізація
- спілкування всередині осередків
- іонний канал-зв'язаний рецептор
- клітинно-поверхневий рецептор, який утворює плазмовий мембранний канал, який відкривається при зв'язуванні ліганду з позаклітинним доменом (ліганд-горовані канали)
- ліганд
- молекула, що виробляється сигнальної клітиною, яка зв'язується зі специфічним рецептором, доставляючи сигнал в процесі
- нейромедіатор
- хімічний ліганд, який несе сигнал від однієї нервової клітини до іншої
- паракринний сигнал
- сигнал між сусідніми клітинами, який доставляється лігандами, що рухаються в рідкому середовищі в просторі між клітинами
- рецептора
- білок в або на клітині-мішені, які зв'язуються з лігандами
- сигнальна комірка
- клітина, яка вивільняє молекули сигналу, що дозволяють спілкуватися з іншою
- синаптичний сигнал
- хімічний сигнал (нейромедіатор), який подорожує між нервовими клітинами
- клітина-мішень
- клітина, яка має рецептор для сигналу або ліганду з сигнальної клітини