6.1: Вступ
- Page ID
- 6552

хлоропласти

Кредит: Ельвінсонг [CC-BY-SA 3.0]
Хлоропласти виникли через другу ендосимбіотичну подію у рослин та різних протестів. Ці органели, що збирають світло, мають схожість за структурою та геномом з фотоавтотрофними ціанобактеріями.
Світло Збирання

Тилакоїдні мембрани хлоропластів і ціанобактерій забезпечують додаткову площу поверхні для енергетичного захоплення світла. Світлозалежні реакції в хлоропластах використовують два білкових комплекси, які називаються фотосистемою I (PSI) та Photosystem II (PSII), розташованими на тилакоїдних мембранах. У центрі кожної фотосистеми комплекси знаходяться фотопігменти, оптимізовані для поглинання конкретних довжин хвиль світла. Коли світло поглинається фотосистемою, електрон збуджується і передається в ланцюг транспорту електронів. У PSII електрон регенерується шляхом розщеплення двох молекул води на 4H + + 4e — + O 2. Коли електрони рухаються через ETC, протони закачуються в тилакоїдний простір. ETC призводить до зменшення високоенергетичного носія електронів NADP + до NADPH. Оскільки цей шлях використовує споживає воду в хімічній реакції, очевидна втрата води в тилакоїдному просторі називається хеміосмосом.
PSI також відомий як циклічний шлях, оскільки збуджений електрон проходить через замкнутий контур ETC для регенерації втраченого електрона. Цей замкнутий контур також генерує градієнт протона за рахунок живлення протонного насоса, але не призводить до зменшення NADPH. Як і у випадку з протоновим насосом, що працює на ETC в мітохондріях, градієнт протона використовується для живлення АТФ-синтази при виробництві молекул АТФ.
Легкі незалежні реакції

Кредит: Майк Джонс [CC-BY-SA 3.0]
Незалежні від світла реакції також відомі як темні реакції або цикл Кальвіна і використовують АТФ і NADPH від світлозалежних реакцій для фіксації газоподібного CO 2 у вуглеводні магістралі. Фотосинтез часто спрощується в 6CO 2 + 6H 2 O + світло —> C 6 H 12 O 6 + 6O 2. Однак справжнім продуктом є 3-фосфогліцерин, який можна використовувати для отримання довших вуглеводів, таких як глюкоза. Відправною точкою фіксації вуглецю є вуглеводна рибулоза 1,5-бісфосфат. Фермент рибулоза бісфосфат карбоксилаза (RubiSCo) захоплює молекулу CO 2 на рибулозу 1,5-бісфосфат, щоб генерувати 2 молекули 3-фосфогліцерату, які можуть вступати в процес глюконеогенезу для генерації глюкози. АТФ від світлових реакцій може сприяти перетворенню 3-фосфогліцерату в 1,3 бісфосфогліцерату, який може бути зменшений NADPH до гліцеральдегід-3-фосфату (G3P). Потім G3P може бути використаний для регенерації рибулози 1,5-бісфосфату.

1: Фіксація вуглецю за допомогою RubiSco

2: Зменшення за NADPH

3: рибулоза, регенерація 5-бісфосфату
Велика подія оксигенації

Дві оцінки еволюції атмосферного O 2. Верхня червона і нижня зелена лінії представляють діапазон оцінок. Стадія 1 (3,85—2,45 Га) являє собою первісну відновлювальну атмосферу. Стадія 2 (2.45—1.85 Ga) збігається з появою океанічних ціанобактерій, де O 2 поглинається океанами та осадами. O 2 врятувався від океанів під час етапу 3 (1,85—0,85 Га). O 2 раковини заповнені в стадії 4 (0,85-0,54 Га) і стадії 5 (0,54 Га - теперішній час), що призводить до накопичення атмосфери.

Смугасті залізні освіти в породі 2,1 мільярда років ілюструють окислення розчиненого океанічного заліза, яке випало в осад у відповідь на накопичення концентрацій O 2.
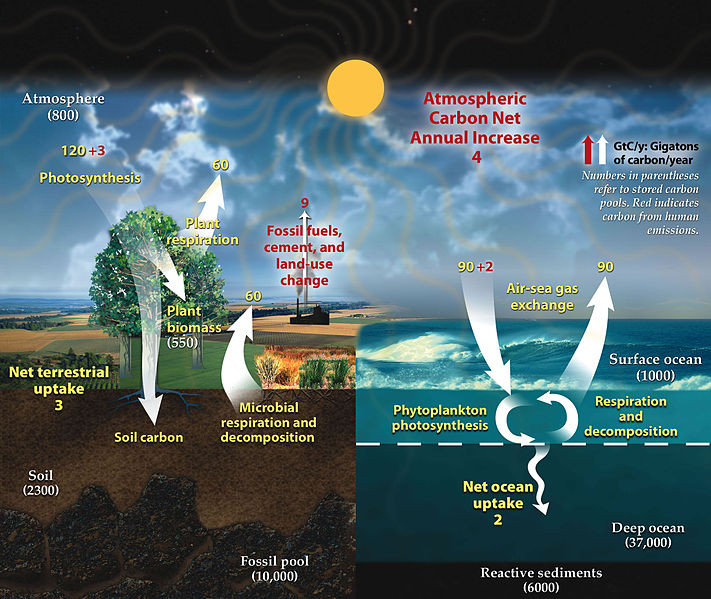
Цикл вуглецю ілюструє секвестрацію вуглецю та вивільнення між різними раковинами вуглецю.

Проекція накопичення атмосферного CO 2 без зменшення скорочення викопного палива NASA.