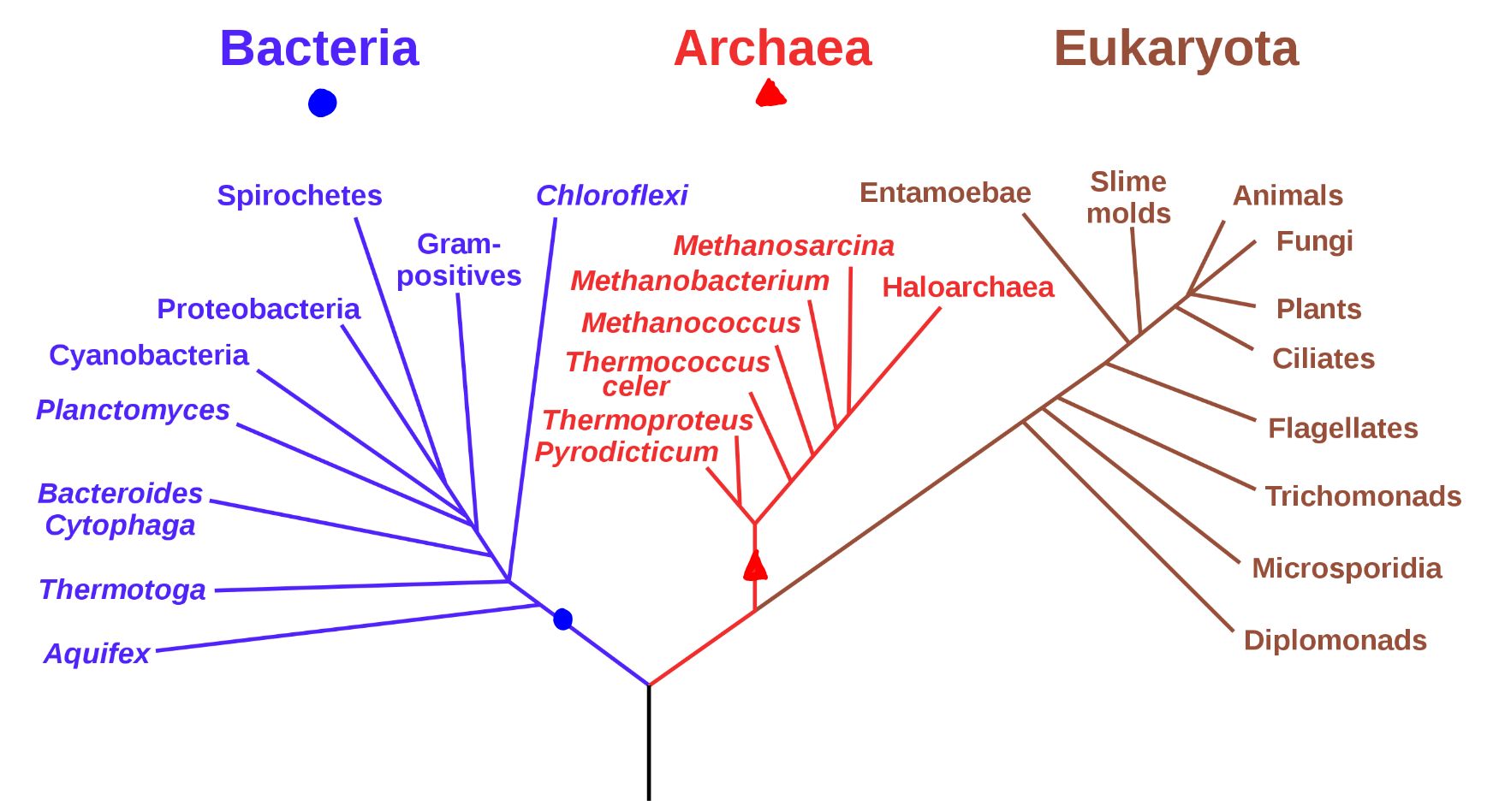
Станом на 1970-ті роки життя було класифіковано на три області: бактерії, архея та Еукарія (показано на малюнку\(\PageIndex{1}\) як Еукаріота). Вважається, що все відоме життя на Землі походить від єдиного спільного предка: ЛУКА (останнього загального загального предка). Взаємозв'язок між трьома областями життя зображена в філогенетичному дереві нижче. Ця діаграма представляє Архею та Еукарію як сестринські таксони, більш тісно пов'язані, ніж будь-яка група з бактеріями домену.
Малюнок\(\PageIndex{1}\): «Філогенетичне дерево живих істот, засноване на даних РНК та запропоноване Карлом Вьесе, що показує поділ бактерій, архей та еукаріотів. Дерева, побудовані з іншими генами, як правило, схожі, хоча вони можуть розміщувати деякі ранньо-розгалужені групи дуже по-різному, завдяки довгому залученню гілок. Точні зв'язки трьох доменів все ще обговорюються, як і положення кореня дерева. Також було висловлено припущення, що через бічну передачу генів дерево може бути не найкращим відображенням генетичних зв'язків усіх організмів. Наприклад, деякі генетичні дані свідчать про те, що еукаріоти еволюціонували внаслідок об'єднання деяких бактерій та архей (одна стає органелою, а інша - основною клітиною)». Ця векторна версія випущена у 2006 році: Ерік Габа (Sting, fr:Sting), Черкаси, Громадське надбання, через Wikimedia Commons.
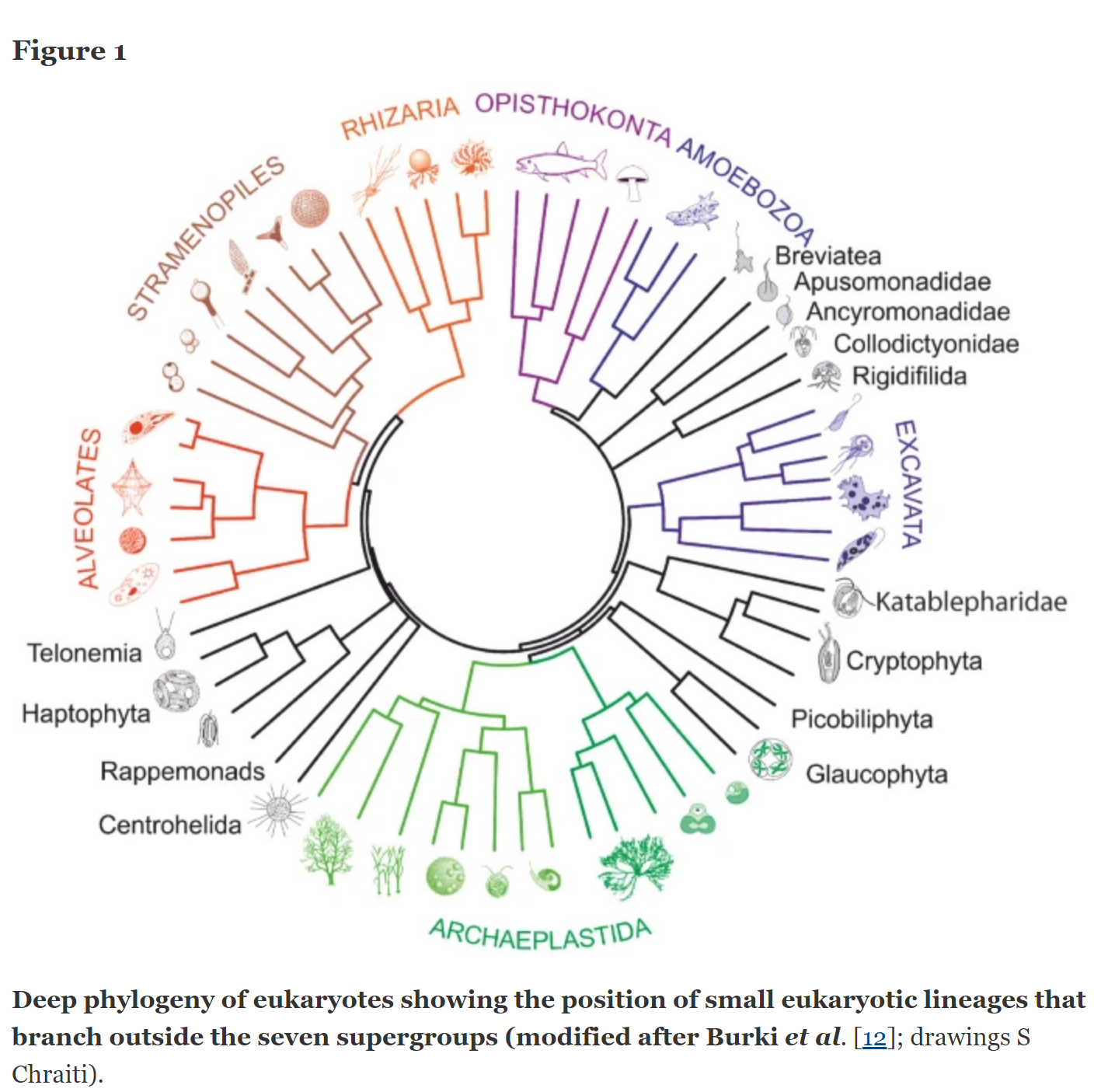
Відносини всередині Еукарії
зображуючи відносини
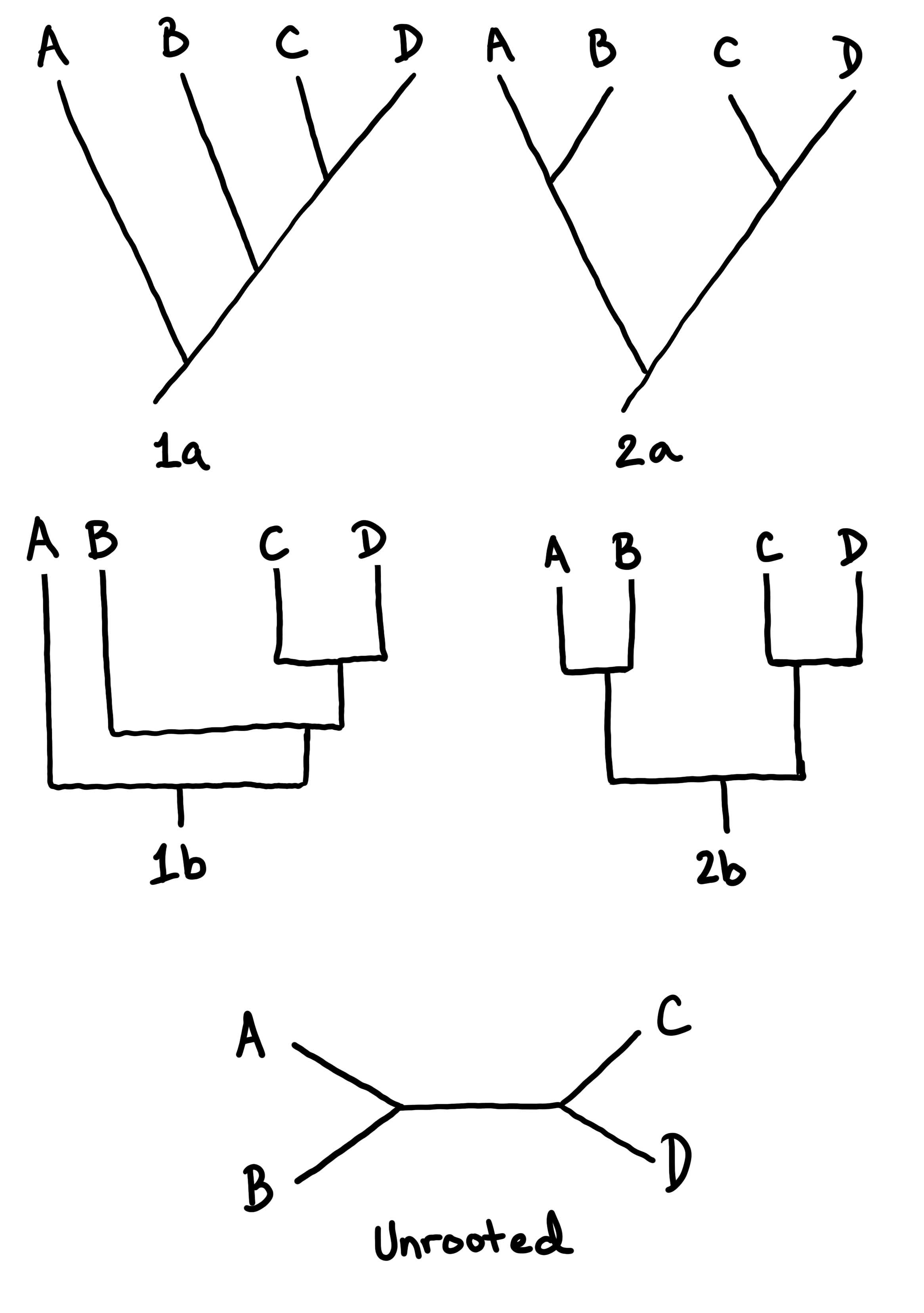
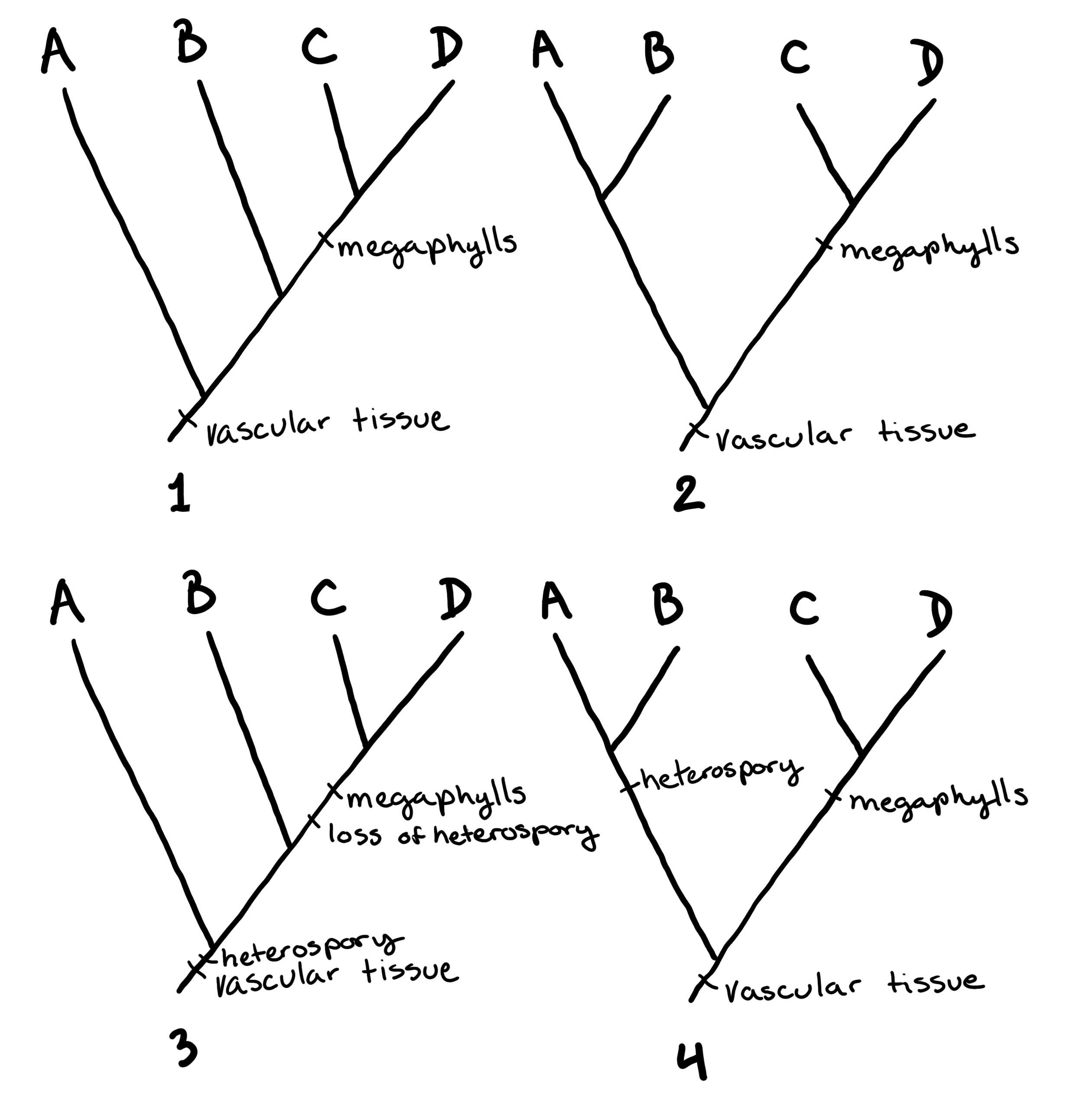
Діаграми гіллястих дерев (як показано вище філогенії) використовуються для передачі гіпотези про те, як організми пов'язані між собою. На малюнку\(\PageIndex{3}\) та малюнку деякі приклади дерев намальовані\(\PageIndex{4}\), щоб показати, як це працює.
Малюнок\(\PageIndex{3}\): Ці діаграми (кладограми) показують гіпотези для чотирьох різних таксонів: A, B, C і D. Ці таксони можуть представляти особин, різні види або будь-який інший рівень класифікації. Дерева 1a і 1b малюються по-різному, але представляють одну і ту ж гіпотезу: C і D найбільш тісно пов'язані (сестринські таксони), а A - найбільш родове походження. Дерева 2a і 2b представляють дещо іншу гіпотезу: C і D - сестринські таксони, але A і B також сестринські таксони; жодна лінія не зображується як «старша» за іншу. Це останнє співвідношення також представлено в дереві «Некорені»: у цьому дереві немає гіпотези, для якої групи є найбільш предковою (немає «кореня»). Зображення, намальовані Марією Морроу, CC-BY.Малюнок\(\PageIndex{4}\): У цих чотирьох деревах ми знову розглядаємо гіпотези про те, як пов'язані A, B, C і D. Однак у цих деревах ми маємо деяку інформацію про те, чому були зроблені ці визначення: були включені риси. У деревах 1 і 2 до двох різних гіпотез, представлених на малюнку, додано судинну тканину і мегафіли\(\PageIndex{3}\). Судинна тканина з'являється на корені дерев як родова риса. Це означає, що A, B, C і D всі мають судинну тканину. Мегафіли з'являються в обох деревах як спільна похідна характеристика (синоморфія) C і D. Це означає, що C і D мають мегафіли, а А і В - ні. І дерево 1, і дерево 2 однаково парсімоніальні. У деревах 3 і 4 додана третя риса: гетероспорія. На додаток до інформації з дерев 1 і 2, ми тепер бачимо, що A і B гетероспористі, в той час як C і D - ні. У дереві 3 гетероспорія з'являється як родова риса, яка втрачається в C і D. У дереві 4 гетероспорія є синапоморфією A і B. Дерево 3 має чотири зміни, тоді як дерево 4 має лише три. Це означає, що дерево 4 є більш парсимонійним і тому більш імовірним (хоча і не обов'язково правильним!). Зображення, намальовані Марією Морроу, CC-BY.
Генетична інформація як риси
Малюнок\(\PageIndex{4}\): На цьому зображенні показані послідовності амінокислот для одного і того ж білка (VEGF) у людей і мишей. Послідовності були вирівняні, щоб знайти подібності та відмінності. Смуги через послідовності показують амінокислоти, які не змінюються у випробуваних організмах. Ці позиції вважаються спільними родовими характеристиками. У всіх інших місцях можуть бути відмінності в амінокислотах між організмами. Ми можемо використовувати їх як синоморфи для побудови дерева, припускаючи, що організми, які поділяють більше подібних послідовностей, більш тісно пов'язані між собою. Ця інформація зробила революцію в тому, як будуються філогенії. «Вирівнювання VEGF гомологічного домену репрезентативних VEGF (всіх ссавців VEGF від людини та миші плюс один VEGF-E (вірусний) та один VEGF-F (зміїна отрута)» Mjeltsch, CC BY-SA, через Wikimedia Commons.