5.3: Видоутворення
- Page ID
- 30123
Біологічне визначення видів, яке працює для статевого розмноження організмів, є групою фактично або потенційно схрещуються особин. Згідно з цим визначенням, один вид відрізняється від іншого можливістю спаровування між особинами від кожного виду для отримання родючого потомства. Є винятки з цього правила. Багато видів досить схожі, що гібридне потомство можливо і часто може зустрічатися в природі, але для більшості видів це правило зазвичай дотримується. Насправді наявність гібридів між подібними видами свідчить про те, що вони, можливо, походять від одного виду схрещування і що процес видоутворення може ще не завершитися.
З огляду на надзвичайне різноманіття життя на планеті повинні бути механізми видоутворення: формування двох видів з одного оригінального виду. Дарвін уявляв цей процес як подію розгалуження і намалював процес на єдиній ілюстрації, знайденій у розділі «Про походження видів» (Малюнок нижче). Щоб видоутворення відбулося, з однієї вихідної популяції повинні бути сформовані дві нові популяції, і вони повинні розвиватися таким чином, щоб особини з двох нових популяцій стали неможливими для схрещування. Біологи запропонували механізми, за допомогою яких це може статися, які діляться на дві широкі категорії. Алопатрична видоутворення, що означає видоутворення в «інших батьківщині», передбачає географічне відокремлення популяцій від батьківського виду та подальшу еволюцію. Симпатрічна видоутворення, що означає видоутворення на «тій же батьківщині», включає видоутворення, що відбувається всередині батьківського виду, залишаючись в одному місці.
Біологи розглядають події видоутворення як розщеплення одного родового виду на два види нащадків. Немає жодної причини, чому не може бути більше двох видів, сформованих одночасно, за винятком того, що це менш імовірно, і такі численні події також можуть бути розглянуті як одиночні розколи, що відбуваються близько в часі.

Географічно безперервна популяція має генофонд, який є відносно однорідним. Потік генів, рух алелів по асортименту виду, є відносно вільним, оскільки особини можуть рухатися, а потім спаровуватися з особинами на новому місці. Таким чином, частота алеля на одному кінці розподілу буде аналогічна частоті алеля на іншому кінці. Коли популяції стають географічно переривчастими, що вільний потік алелів запобігається. Коли це поділ триває протягом певного періоду часу, дві популяції здатні розвиватися за різними траєкторіями. Таким чином, їх алельні частоти на численних генетичних локусах поступово стають все більш різними, оскільки нові алелі самостійно виникають шляхом мутації в кожній популяції. Як правило, умови навколишнього середовища, такі як клімат, ресурси, хижаки та конкуренти, для двох популяцій будуть відрізнятися, що спричиняє природний відбір сприяти різним адаптаціям у кожній групі. Різні історії генетичного дрейфу, посилені, оскільки популяції менші, ніж материнська популяція, також призведе до розбіжності.
Враховуючи достатньо часу, генетична та фенотипічна розбіжність між популяціями, швидше за все, вплине на характери, які впливають на розмноження настільки, що були особини двох популяцій, спарювання було б менш імовірним, або якщо відбулося спаровування, потомство було б нежиттєздатним або безплідним. Багато типів розходяться характерів можуть впливати на репродуктивну ізоляцію (неможливість схрещуватися) двох популяцій. Ці механізми репродуктивної ізоляції можна розділити на презиготичні механізми (ті, що діють до запліднення) і постзиготичні механізми (ті, що діють після запліднення). Презиготичні механізми включають риси, які дозволяють особинам знаходити один одного, такі як терміни спарювання, чутливість до феромонів або вибір місць спарювання. Якщо люди здатні зіткнутися один з одним, розбіжність характеру може перешкодити ритуалам залицяння привести до спаровування або тому, що жіночі уподобання змінилися або змінилася чоловіча поведінка. Фізіологічні зміни можуть перешкодити успішному заплідненню, якщо може відбутися спарювання. Постзиготичні механізми включають генетичні несумісності, які перешкоджають правильному розвитку потомства, або якщо потомство живе, вони можуть бути не в змозі самостійно виробляти життєздатні гамети, як на прикладі мула, безплідного потомства самки коня і самця осла.
Якщо дві ізольовані популяції повертаються разом, а гібридне потомство, яке утворилося в результаті спаровування між особинами двох популяцій, має меншу виживаність або знижену фертильність, то відбір сприятиме особам, здатним розрізняти потенційних супутників власної популяції. та інше населення. Цей вибір посилить репродуктивну ізоляцію.
Ізоляція популяцій, що призводять до алопатричного видоутворення, може відбуватися різними способами: від річки, що утворює нову гілку, ерозії, що утворює нову долину, або групи організмів, що подорожують до нового місця без можливості повернутися, наприклад, насіння, що плавають над океаном, на острів. Характер географічного поділу, необхідного для виділення популяцій, повністю залежить від біології організму та його потенціалу для розгону. Якщо дві популяції літаючих комах проживали в окремих довколишніх долині, є ймовірність, що особини з кожної популяції будуть літати туди-сюди, продовжуючи потік генів. Однак, якби дві популяції гризунів розділилися утворенням нового озера, продовження потоку генів було б малоймовірним; отже, видоутворення було б більш імовірним.
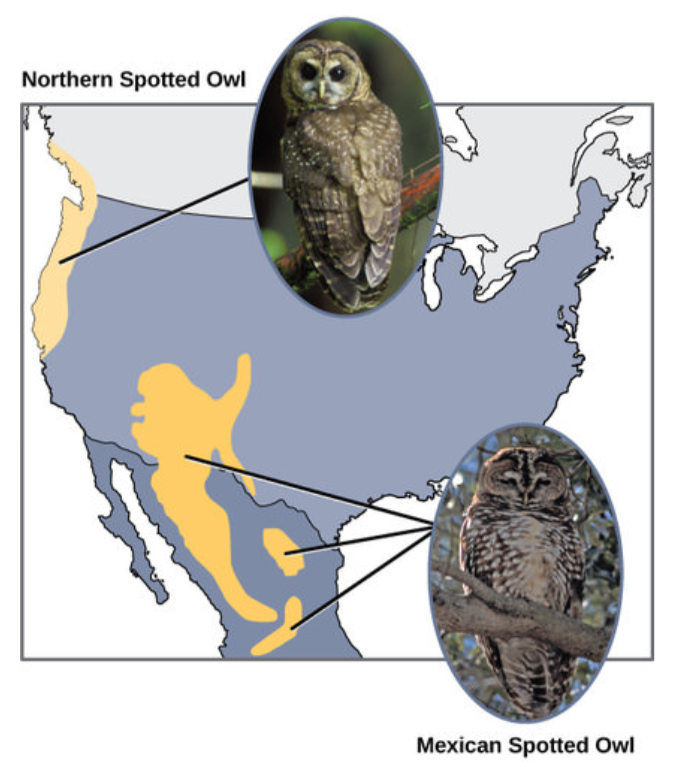
Біологи групують алопатричні процеси на дві категорії. Якщо кілька представників виду переїжджають в нову географічну зону, це називається розгоном. Якщо виникає природна ситуація для фізичного поділу організмів, це називається вікаріансом. Вчені задокументували численні випадки алопатричного видоутворення. Наприклад, уздовж західного узбережжя США існують два окремих підвиди плямистих сов. Північна плямиста сова має генетичні та фенотипічні відмінності від свого близького родича, мексиканської плямистої сови, яка мешкає на півдні (рисунок нижче). Причина їх початкового поділу не ясна, але це, можливо, було викликано льодовиками льодовикового періоду, що розділяють початкову популяцію на дві частини.
У деяких випадках популяція одного виду розходиться по всій території, і кожен знаходить окрему нішу або ізольоване середовище проживання. З часом різноманітні вимоги їх нового способу життя призводять до численних подій видоутворення, що походять від одного виду, який називається адаптивним випромінюванням. З однієї точки походження багато адаптацій розвиваються, змушуючи вид випромінювати кілька нових. Островні архіпелаги, такі як Гавайські острови, забезпечують ідеальний контекст для адаптивних радіаційних подій, оскільки вода оточує кожен острів, що призводить до географічної ізоляції для багатьох організмів (рисунок нижче). Гавайський медоносник ілюструє один приклад адаптивного випромінювання. З одного виду, званого видом-засновником, еволюціонували численні види, включаючи вісім, показаних на малюнку нижче.
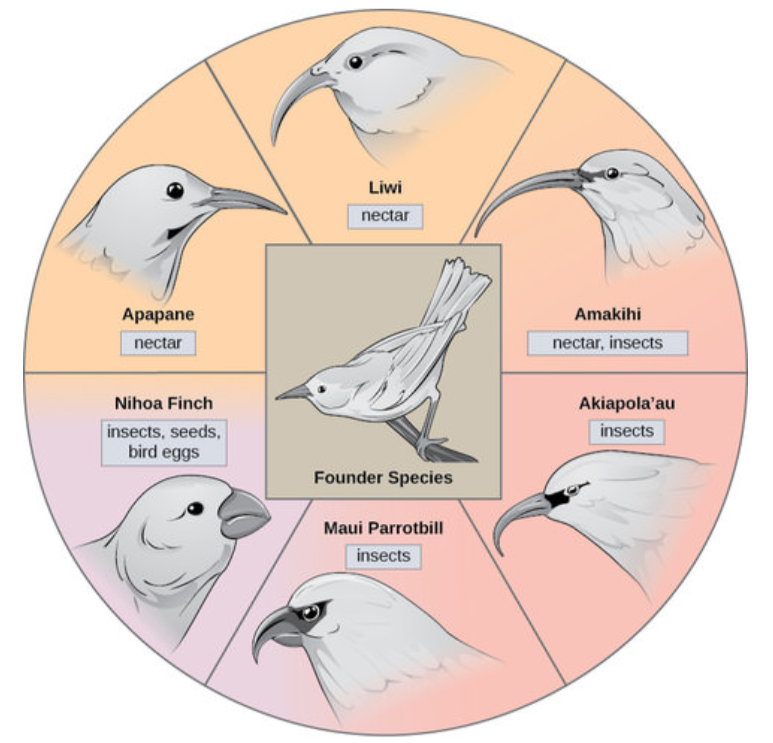
Птахи медоносні ілюструють адаптивне випромінювання. Від одного оригінального виду птахів еволюціонувало безліч інших, кожен зі своїми відмінними характеристиками. Зверніть увагу на відмінності дзьобів виду на малюнку вище. Зміна генетичної варіації дзьобів у відповідь на природний відбір на основі конкретних джерел їжі в кожному новому середовищі існування призвела до еволюції іншого дзьоба, придатного до конкретного джерела їжі. Птахи, що їдять плоди та насіння, мають більш товсті, міцніші дзьоби, які підходять для розбивання твердих горіхів. Птахи, що їдять нектар, мають довгі дзьоби, щоб зануритися в квіти, щоб досягти свого нектару. Птахи, що їдять комах, мають дзьоби, як мечі, придатні для колючих і насаджування комах. Дарвінові в'юрки - ще один добре вивчений приклад адаптивного випромінювання на архіпелазі.
Видоутворення без географічного поділу
Чи може статися розбіжність, якщо немає фізичних бар'єрів для відокремлення особин, які продовжують жити і розмножуватися в одному середовищі існування? Запропоновано та вивчено низку механізмів симпатрічного видоутворення.
Одна форма симпатрічного видоутворення може починатися з хромосомної помилки під час мейозу або формування гібридної особини з занадто великою кількістю хромосом. Поліплоїдія - це стан, при якому клітина, або організм, має додатковий набір, або набори хромосом. Вчені виділили два основних види поліплоїдії, які можуть привести до репродуктивної ізоляції індивіда в поліплоїдному стані. У деяких випадках поліплоїдна особина матиме два або більше повних наборів хромосом від власного виду в стані, званому автополіплоїдією. Приставка «авто» означає себе, тому термін означає множинні хромосоми від власного виду. Поліплоїдія виникає в результаті помилки мейозу, при якій всі хромосоми переміщаються в одну клітину замість відокремлення.
Наприклад, якщо вид рослин з 2 n = 6 виробляє аутополіплоїдні гамети, які також є диплоїдними (2 n = 6, коли вони повинні бути n = 3), то гамети тепер мають вдвічі більше хромосом, ніж вони повинні мати. Ці нові гамети будуть несумісні зі звичайними гаметами, виробленими цим видом рослин. Але вони могли або самозапилюватися, або розмножуватися іншими автополіплоїдними рослинами з гаметами, що мають однакове диплоїдне число. Таким чином, симпатрічне видоутворення може відбуватися швидко, утворюючи потомство з 4 n, званим тетраплоїдом. Ці особини негайно зможуть розмножуватися лише з тими, хто має цей новий вид, а не з родовими видами. Інша форма поліплоїдії виникає, коли особини двох різних видів розмножуються, утворюючи життєздатне потомство, яке називається алополіплоїдом. Приставка «алло» означає «інший» (нагадаємо з аллопатрического); тому алополіплоїд виникає при поєднанні гамет двох різних видів.
Культурні форми рослин пшениці, бавовни та тютюну - все це алополіплоїди. Хоча поліплоїдія трапляється зрідка у тварин, більшість хромосомних аномалій у тварин смертельні; це відбувається найчастіше у рослин. Вчені виявили, що більше 1/2 всіх досліджуваних видів рослин відносяться до виду, що розвинувся за допомогою поліплоїдії.
Симпатрічна видоутворення може також відбуватися іншими способами, ніж поліплоїдія. Наприклад, уявіть собі вид риб, який мешкав в озері. У міру зростання населення зростала і конкуренція за продукти харчування. Під тиском, щоб знайти їжу, припустимо, що група цих риб мала генетичну гнучкість, щоб виявити і годувати інший ресурс, який був невикористаний іншою рибою. Що робити, якщо це нове джерело їжі було знайдено на іншій глибині озера? З часом ті, хто харчується другим джерелом їжі, більше взаємодіятимуть між собою, ніж інші риби; тому вони також розмножуватимуться разом. Потомство цих риб, швидше за все, поводитиметься як їхні батьки і годуватиме та живе в одній місцевості, зберігаючи їх окремо від початкової популяції. Якщо ця група риб продовжувала залишатися відокремленою від першої популяції, врешті-решт симпатичне видоутворення може виникнути, оскільки між ними накопичується більше генетичних відмінностей.
Цей сценарій дійсно грає в природі, як і інші, які призводять до репродуктивної ізоляції. Одним з таких місць є озеро Вікторія в Африці, відоме своїм симпатичним видом цихлідних риб. Дослідники виявили сотні симпатрічних подій видоутворення у цих риб, які відбулися не тільки у великій кількості, але і за короткий проміжок часу. На малюнку нижче показаний цей вид серед популяції цихлід риби в Нікарагуа. У цій місцевості два типи цихлід живуть в одному географічному положенні; однак вони мають різну морфологію, яка дозволяє їм їсти різні джерела їжі.

Нарешті, добре задокументований приклад триваючого симпатрічного видоутворення стався у яблучної мухи опариша, Rhagoletis pomonella, яка виникла як ізольована популяція через деякий час після введення яблука в Північну Америку. Рідна популяція мух харчується видами глоду і є специфічною для господаря: вона заражає тільки дерева глоду. Важливо, що він також використовує дерева як місце для зустрічі для спарювання. Викладається гіпотеза, що або через мутацію, або поведінкову помилку, мухи стрибали господарів і зустрічалися і спаровувалися на яблунях, згодом відкладаючи свої ікри в плоди яблуні. Потомство дозріло і зберігало перевагу яблуням, ефективно розділяючи початкову популяцію на дві нові популяції, розділені видами господарів, а не географією. Стрибок господаря відбувся в дев'ятнадцятому столітті, але зараз існують вимірні відмінності між двома популяціями мух. Здається ймовірним, що специфіка паразитів хазяїна взагалі є поширеною причиною симпатрічного видоутворення.