20.19: Зберігання інформації
- Page ID
- 23355

Як молекули ДНК та РНК можуть діяти як креслення для виготовлення білків? Точні деталі були розгадані на початку 1960-х в основному Маршаллом Ніренбергом (нар. 1927) в Національних інститутах охорони здоров'я і H.G. Khorana (нар. 1922) в Університеті Вісконсіна, робота, яка принесла їм Нобелівську премію в 1968 році. Вони показали, що кожна амінокислота в білку визначається специфічним кодоном з трьох азотистих основ в ланцюжку ДНК або РНК. Подробиці цього генетичного коду наведені в таблиці нижче. Як приклад того, як працює цей код, візьмемо розділ РНК, показаний на малюнку 3, про структуру нуклеїнових кислот. Це має послідовність UCAUGG. Це частина інструкції з побудови поліпептидного ланцюга, що містить амінокислоту серин (UCA), за якою слідує амінокислота триптофан (UGG).
\(\PageIndex{1}\)Таблиця Генетичний код для РНК
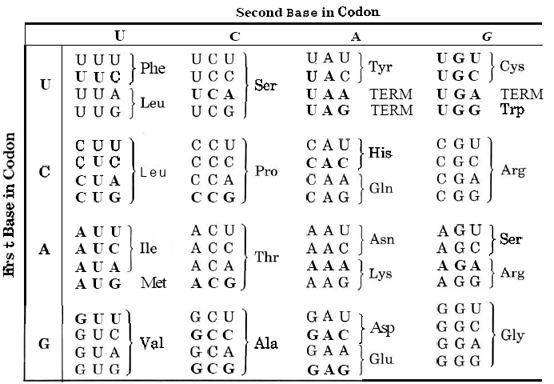
(а) Кодон припинення позначається ТЕРМІНОМ. (b) AUG, кодон для метіоніну також є кодоном ініціації. Весь синтез білка починається з цього кодону, хоча цей початковий метіонін часто видаляється під час посттранскрипційної обробки.
Існує три додаткові особливості генетичного коду. По-перше, AUG, кодон для метіоніну також служить кодоном ініціації, і за допомогою інших сигналів починається синтез білка. Друга особливість полягає в тому, що читання РНК для синтезу білка йде від 5-дюймового вуглецевого кінця нуклеїнової кислоти до 3-футового вуглецевого кінця. Остаточною важливою особливістю генетичного коду є існування трьох кінцевих кодонів. Вони відповідають інструкції по закінченню поліпептидного ланцюга. Як працюють ці функції найкраще проілюструвати на прикладі.
Розшифрувати фрагмент РНК
5' А С С У У А У Г А С Г С С У Г У С У С А У У А А С Г А У 3'
Рішення
По-перше, ми повинні вирішити, в якому напрямку читати код РНК. Синтез йде від кінця 5' до кінця 3', так що цей сегмент читається зліва направо. Якби він відображався від 3 'до 5', нам потрібно було б прочитати його справа наліво.
По-друге, нам потрібно шукати кодон ініціації, AUG. Цей кодон з'являється починаючи з шостої літери в. Таким чином, ми можемо розділити послідовність вгору так, з початковим кодоном жирним шрифтом:
- AC|CUU| СЕРПНЯ |ACG|CCU|GUC|КАУ|UAA|CGA|U
По-третє, давайте подивимося, чи є стоп-кодон в цій послідовності. Звичайно, п'ятий кодон після стартового кодону, UAA - це стоп-кодон. Таким чином, вся послідовність, яку потрібно перевести, виділено жирним шрифтом:
- AC|CUU| АУГ|АКГ|ККУ|ГУК|КАУ|УАА |КГА|У
що перекладається на амінокислотну послідовність:
- Мет-THR-про-валь-хіс-стоп
Зауважте в прикладі, що якби ми не почали з кодоном ініціації, утворився б зовсім інший білок. Подивіться, що б сталося, якби ми просто почали на початку послідовності:
- АКК|УУА|УГА |КГК|КГ|УКК|АУУ|ААА|ГАУ|
на новому місці з'являється стоп-кодон, а перекладається білок - це:
- THR-Леу-стоп
Це підкреслює важливість кадру читання, місця, де кодони починають читатися. Зверніть увагу, що, оскільки кодони мають довжину 3 основи, будь-яка послідовність має три різні кадри читання. Без кодону ініціації не було б можливості визначити правильний кадр читання. Крім кодону ініціації AUG, інші елементи регулюють ініціацію. У бактерій послідовність основ перед кодоном ініціації, звана послідовністю Shine-Dalgarno, передує кодону AUG, вказуючи, з чого почати переклад. Інша установка відбувається у еукаріотів. Формується комплекс ініціації, але замість того, щоб мати певну послідовність, з'єднану з кодоном ініціації, комплекс ковзає по ланцюжку мРНК, поки не знайде кодон ініціації AUG. [2]
- ↑ Нельсон, Д.Л., Кокс, М.М.Ленінгер Принципи біохімії (<sup>5-е</sup> видання). Нью-Йорк: W.H Фрімен і компанія, 2008. стор. 1070-1072.
- ↑ Нельсон, Д.Л., Кокс, М.М.Ленінгер Принципи біохімії (<sup>5-е</sup> видання). Нью-Йорк: W.H Фрімен і компанія, 2008. стор. 1088-1090.