1.6: Ранній розвиток
- Page ID
- 68201

- Огляд раннього розвитку
- Візерунки
- Перший триместр:
- Передімплантація
- Гаструляція
- Диференціація ектодерми
- Нейруляція
- Міграція нейронного гребеня
- Диференціація мезодерми
- утворення сомітів
- Формування серця
- Диференціація ентодерми
- Формування глотки
- Розвиток інших структур порожнини рота
- Розвиток слинних залоз
- Розвиток мовних сосочків
- Клінічні застосування раннього розвитку

Огляд раннього розвитку
Розвиток людини - це процес, коли одна клітина, візуально нічим не примітна клітина, крім хіба що її розміру, повільно змінюється на трильйони (37) різних на вигляд клітин. Ембріологія - це вивчення фаз розвитку, особливо ранніх фаз, де відбувається більшість цікавого. Кругла клітина, просто кулька без боків і без особливих частин, вростає в організм з різними органами в різних місцях. Рис. 6.1 - Сонце. Ви бачите це кожен день, чи можете ви сказати, чи є фотографія правою стороною вгору? Чи є навіть права сторона вгору? Ми не астрономи, але Сонце трохи схоже на запліднену яйцеклітину, і на рис. 6.1 немає явних орієнтирів для орієнтації нашого погляду. Яйцеклітина має схожий аморфний (без форми) вигляд, але воно морфно (плавно змінює форму) в ембріон, показаний на рис. 6.2, який має головку і хвостовий кінець, і чіткі початку рук і ніг. Перша частина цього розділу зосереджена на поняттях, загальних для багатьох подій розвитку, що спостерігаються в главах 7-11. Друга половина глави охоплює деталі раннього розвитку.
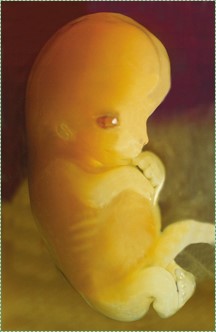
Вагітність ділиться на три рівні часові періоди, названі триместрами. Протягом першого триместру одна клітина розмножується в багатоклітинний організм без помітної форми (крім круглої краплі, а не ікозоедра). Починаючи з 2 тижня, пляма клітин переростає в ембріон, який має різну форму. Ембріон продовжує розвивати нові форми, поки не матиме всіх основних людських форм, таких як голова, руки та серце. У цей час (8 тиждень) це плід, і закінчився перший триместр. Наступні два триместри відносно нудні, плід зростає більше, а форми дозрівають, але нічого жахливо захоплюючого не відбувається до народження. Ось чому це книга з гістології та ембріології, а не гістології та фетології.


Оскільки клітини людини діляться протягом першого триместру, вони приймають рішення щодо своєї клітинної долі, що в кінцевому підсумку визначає кількість і розташування наших рук і ніг, нашої голови та серця. Ці рішення передбачають сигнали, що надсилаються між клітинами. Помилки в цих сигналах або рішеннях призводять до вроджених порушень (або вродженої вади розвитку, або вроджених дефектів (але див. Нижче)). Це може статися без відомої причини. Або хімічні речовини можуть перешкоджати сигналам розвитку, і ми називаємо ці хімічні речовини тератогенами. Найвідомішим тератогеном у моїй країні є препарат талідомід, який - якщо приймати вагітні жінки в неправильний час - втручається в процеси прийняття рішень, які ембріон використовує, щоб вирішити, де рости кінцівки. Це викликає фокомелію, або мальформацію кінцівок. Не кожна країна поділяє цю упередженість, тератогенні ефекти агента апельсина (діоксину, який також міститься в сигаретному димі) та атомної радіації (pdf download) помітно відсутні в більшості американських шкільних класів (агент оранжевого та атомного випромінювання також мають несприятливий вплив на дорослих, і швидкість, яку ця інформація доходить до американської громадськості, схильна до подібних упереджень). Ми розглянемо невелику кількість сигналів, які беруть участь у деяких важливих рішеннях, які приймаються під час розвитку обличчя та зубів. 37 трильйонів рішень приймається між одноклітинною стадією і пологами, помилки неминучі. Деякі помилки незначні, змінюючи зовнішній вигляд, але не функцію органу; ми можемо називати це як знак народження або зміна людини. Інші помилки є більш серйозними, заважають здоров'ю людини, і ми називаємо їх вродженими порушеннями (або, можливо, вродженими помилками розвитку, але бажано не вродженими дефектами, оскільки наш вибір слів може впливати на кількість медичної допомоги, яку люди шукають, це залежить від пацієнта). Якщо ви в будь-який час відчуваєте, що вам потрібна додаткова інформація про ембріологію, ці посилання корисні:
- Сторінка «Освіта ембріології»
- доктор Марк Хілл в Університеті Нового Південного Уельсу, Сідней, Австралія
- Зображення ембріонів Нормальний і ненормальний розвиток ссавців
- д-р Кетлін К. Сулік та д-р Пітер Лям-молодший в Медичній школі Університету Північної Кароліни.
- 3D атлас ембріології людини
- колекція 3D-файлів PDF з відкритим вихідним кодом (читається за допомогою Adobe Acrobat reader) понад 70 різними студентами Академічного медичного центру, Нідерланди.
Візерунки
У цьому розділі розглядаються основні процеси, що відбуваються під час розробки. Найчастіше в одному і тому ж місці одночасно відбувається не один процес. Тим не менш, корисно думати про наступні процеси окремо, як ви могли б зосередитися тільки на vocal— або віолончелі, або гітарі, або accordion— в пісні.

Клонування
Мітоз виробляє клони, або ідентичні клітини з ідентичною ДНК. Однак клітини диференціюють і експресують різні гени, щоб стати різними тканинами та органами. У перші 2 тижні розвитку клітини діляться і виробляють більше однакових (хоча і менших) клітин. Дуже скоро клітини приймають рішення про те, якими вони збираються стати, і ці рішення узгоджуються з іншими клітинами. Без координації ембріон міг би закінчитися двома недопалками і без голови.

Диференціація
Диференціація - це процес, коли стовбурові клітини починають експресувати різні гени, змушуючи їх з'являтися і функціонувати інакше, ніж інші клітини. Всі клітини людського організму мають однакову ДНК, тому що всі клітини копіювали свою ДНК з ДНК в зиготу (кілька виняткових клітин трохи змінюють свою ДНК). Зигота є тоти-потужною стовбуровою клітиною, фактори транскрипції можуть потенційно зв'язуватися та активувати будь-який ген. Однак, коли клітини диференціюються, вони обмежують свою клітинну долю, поміщаючи деякі гени в глибоке зберігання (фактично не змінюючи генетичний код, але змінюючи те, що може бути доступним або не може бути доступним для РНК-полімерази).

Диференціація спрацьовує при прийомі хімічних сигналів. Ці сигнали змушують клітини-мішені метилювати ДНК певних непотрібних генів, упаковувати ці ділянки ДНК навколо гістонів і назавжди закривати ці гени. Наприклад, клітини, яким судилося стати яєчниками, ніколи не синтезують інсулін, ген інсуліну упаковується задовго до того, як яєчники виділяють естроген або овуляцію. Метилювання ДНК і упаковка гістонів обмежують долю цих клітин. Після цього сигнали можуть зникнути, оскільки картина метилювання копіюється під час мітозу і передається обом дочірнім клітинам (і їх дочірнім клітинам). На рис. 6.7, уявіть, що сигнал доручає одній клітині метилювати зелені гени, залишаючи червоні гени відкритими для потенційно транскрипції. Інший сигнал робить протилежне іншому осередку. Оскільки еритроцити дублюють свою ДНК, вона копіює схему метилювання, так що всі її дочірні клітини не можуть експресувати зелені гени. Тим часом зелена клітина виробляє ряд дочірніх клітин, які мають червоний ген метильований, жодна з дочірніх клітин не може експресувати червоний ген. Тепер у нас є 4 еритроцити, які належать до одного роду, і 4 зелені клітини, які належать до іншої лінії. Подальше малювання виникає, коли клітини транскрибують неметильовані гени. У мультфільмі, коли червоні клітини транскрибують червоний ген, це створює сигнал короткого діапазону, який спонукає сусідні зелені клітини транскрибувати темно-зелений ген. Ми б сказали, що ці клітини-мішені диференціювалися далі, і цей організм став ще складнішим (3 різні типи клітин). Це повинно тривати, поки у нас не буде 200 або близько того різних типів клітин. Взагалі кажучи, оскільки клітина стає більш спеціалізованою, її доля стає більш обмеженою (більше метилювання ДНК та упаковка гістонів), можливо, до того, що вона обмежена лише долею однієї клітини.

Індукційні
Одна клітина може сигналізувати іншій клітині і спонукати (наказує) цю клітину диференціювати. Хімічні речовини, які можуть спонукати іншу клітину змінювати свою форму (перетворювати) та змінювати долю клітин, називаються морфогенами (або іноді називають факторами росту). Морфогени схожі на нейромедіатори або гормони, вони часто зв'язуються з білками рецепторів поверхні клітин. Активація морфогенних рецепторів призводить до активації внутрішньоклітинних сигналів, які, в свою чергу, можуть або активувати, або пригнічувати специфічні фактори транскрипції для зміни експресії різних генів. Зміни в експресії генів змушують клітину-мішень виглядати і функціонувати по-різному. Морфогени також змінюють метилювання генів та їх упаковку навколо гістонів, що є більш постійною зміною експресії генів. Ця закономірність генної регуляції передається дочірнім клітинам, який ми дізналися в главі 1, є прикладом епігенетичного успадкування. Регулюючи експресію генів через активність фактора транскрипції та метилювання, морфогени змінюють не тільки морфологію клітини-мішені, але і їх клітинну долю. Часто існує кілька тісно пов'язаних молекул, які належать до сімейства морфогенів, кожна хімічна речовина має таку назву, як морфоген-1, морфоген-2а, морфоген-2b та морфоген-3. Існує дуже велика кількість різних морфогенів, у цьому підручнику ми обговорюємо лише деякі за назвою, в основному ті, які безпосередньо пов'язані з розвитком зубів, включаючи сімейство білка морфогену кістки (BMP), сімейство Wnt та сімейство Фактор росту фібробластів (FGF). Морфогени можуть діяти на різних відстанях, перерахованих в таблиці 6.1. Назви конкретних морфогенів дуже важливі на ембріологічних іспитах, тим більше на іспитах з гігієни зубів.
| Відстань | Тип | Приклади |
|---|---|---|
| Пряма | Клітинка-клітинка | Епітеліал-нейро-мезенхімальні стовбурові клітини (Диференціація одонтобласта) |
| Клітина до ECM | Інтегран-колаген (диференціація амелобластів) | |
| Короткий радіус дії | Дифузія через подрібнену речовину | Кістковий морфогенний білок (BMP), Wnt, фактор росту фібробластів (FGF). (Індукція зубного бруньки) |
| Широкі сигнали тіла | Гормони | Гормон росту, кальцитонін. |
Таблиця 6.1: Приклади різного типу сигналів, помічених під час ембріогенезу.
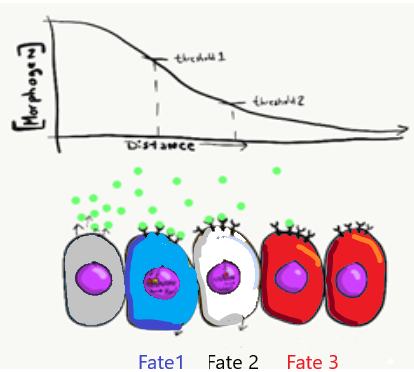
Морфогени виділяються в подрібнену речовину, створюючи градієнт концентрації рівнів морфогену, оскільки вони повільно дифундують від джерела. Кількість вироблених молекул морфогену, відносна липкість подрібненої речовини, відстань від джерела і кількість рецепторних білків на клітині-мішені визначають рівень сигналу, який отримує клітини. Іноді морфоген створює видимі ефекти, які падають уздовж градієнта (або спектру), який концептуально відповідає градієнтам, які ми щойно перерахували, наприклад, колір шкіри (світліший до темніший) або розмір тіла (коротший до вищого). Однак іноді ефекти морфогену більш дискретні (зубна нирка або відсутність зубної нирки, немає половини зубних бруньок). Щоб допомогти пояснити ці спостереження, потрібно трохи знань про те, як працюють фактори транскрипції. Наприклад, у моделі французького прапора (рис. 141) уявіть, що отримання високого рівня морфогенного сигналу запускає транскрипцію синього гена і пригнічує червоний ген, змушуючи ці клітини диференціюватися на сині клітини. Помірної кількості сигналу морфогену може бути недостатньо для набору РНК-полімерази до синього гена, але достатньо для інгібування червоного гена, змушуючи клітини в регіонах помірного рівня морфогену диференціюватися на білі клітини. Нарешті, клітини, які отримують мало або взагалі не отримують морфогенного сигналу, не пригнічують червоний ген, змушуючи їх диференціюватися на еритроцити. Є й інші моделі, які допомагають пояснити зв'язок між індукцією та морфогенезом - між хімічною речовиною та організмом - наприклад, морфогенетичне поле менш франкофільне за своєю природою, але важче візуалізувати. Всі ці моделі поділяють важливу концепцію: хімічні сигнали можуть передавати позиційну інформацію у ембріона, що розвивається.
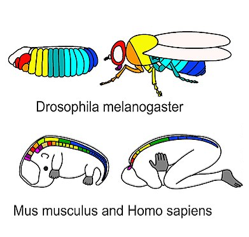
Під час дуже раннього розвитку регіональна експресія генів гомеобоса контролює долю клітин, а не морфологію. Гени Homeobox - це фактори транскрипції, які визначають, до якої частини тіла зараз входить певна клітина, наприклад, частина руки або обличчя, навіть якщо ця область ще не має певної форми. Вони не кодують білки, змушують клітини стати частиною певного органу. Натомість гени гомеобоксу активують або інгібують колекцію інших генів (програму), створюючи основу для майбутніх змін. На відміну від цього, морфогени, такі як FGF та BMP, активують гени, які змінюють форму клітини або тканини (морфологію). Експресія генів Homeobox являє собою дуже специфічний план в ембріології. Він високо зберігається серед видів тварин, і картина дуже передбачувана у ембріонів. Візерунок також відносно простий і лінійний (рис.6.10). Подумайте про гени гомеоблоків, як геодезисти, які визначають місця, де будинки будуть побудовані в підрозділі, тоді як морфогени - це підрядники, які слідують за кресленнями ДНК для кожного будинку, які побудовані клітинами. Ми можемо розширити цю метафору на крок далі: у підрозділі будинки схожі, похідні від одного основного плану, але з урахуванням незначних модифікацій з кожним повторним використанням.
![[анімувати вихідне зображення]](https://med.libretexts.org/@api/deki/files/22913/animate-output-image.gif)
Здатність тканини створювати власні закономірності називається самоорганізацією (навпаки, гени материнського ефекту з клітин мами направляють закономірності в клітині ембріона). Це включає в себе множинні морфогени, спочатку виражені випадковим чином. Взаємодія між цими морфогенами призводить до стабільної картини експресії та складної картини диференціації. Наприклад, деякі морфогени блокують інші клітини виробляти той самий морфоген. На рис. 6.11 уявіть, що синій морфоген спонукає одну клітину диференціюватися в блакитну клітину, але блокує сусідні клітини від перетворення в сині клітини. Ці сусіди могли б вільно диференціюватися на еритроцити. У той же час віддалена клітина виробляє червоний морфоген, який спонукає її диференціюватися в еритроциту, але блокує сусідам робити те саме. Його сусіди могли б вільно виробляти синій морфоген і диференціюватися на сині клітини. Виникає початковий візерунок - це чергування кілець червоних і синіх клітин. Однак кільця утворюють регіони червоно-поруч із червоним та синьо-поруч із синім, що порушує наші правила переконання, і це починає бій. Щоб мінімізувати кількість червоно-поруч з червоним і синьо-поруч з синім, клітини змінюють свою клітинну долю до тих пір, поки не виникне стійкий малюнок, в даному випадку смуги. З самого початку було б важко передбачити, які клітини будуть червоними проти синіх, але можна було б передбачити розвиток червоних і синіх смуг. Якби це були клітини, що виробляють пігмент у зебрі, у нас була б зебра з смужками (найкращий тип зебри). Але ця картина також може бути чергуванням ділянок слизової оболонки порожнини рота, які ростуть зубну бруньку або не ростуть зубну нирку, забезпечуючи, щоб кожна зубна брунька мала достатньо місця для росту (якщо ви хочете спуститися вниз кролячою діркою, ці взаємодії можна моделювати математично). Коли морфоген блокує інший сигнал, ми називаємо цей антагонізм, а сигнали антагоністів так само поширені, як і індукційні сигнали. Вони повинні, врешті-решт, принципово так само важливо відростити одну голову, як і не виростити дві головки.
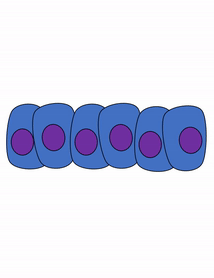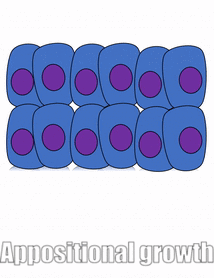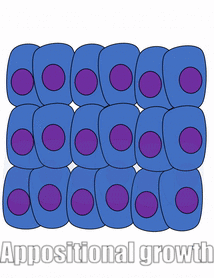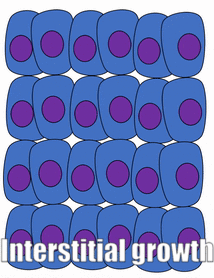
Проліферація
Розростання тканини називається проліферацією. Проліферація відбувається або шляхом мітозу (додавання клітин), або шляхом додавання позаклітинного матриксу (синтезуючи білки або інші молекули). Коли нове зростання відбувається поруч із областю старшого зростання, це апозиційний ріст, тоді як якщо зростання відбувається зсередини і виштовхує старі тканини назовні, тобто інтерстиціальний ріст. Щільні тканини, такі як кістка, емаль і дентин, піддаються апозиціонному зростанню. Більш м'які тканини, такі як мезенхіма, ростуть інтерстиціально.
Зростання тканини регулюється гормонами, такими як гормон росту, який координує швидкість росту тканин по всьому тілу. З іншого боку, морфогени місцевої дії змушують невеликі регіони рости швидше, ніж сусідні регіони. Щоб звільнити місце для нових клітин, зберігаючи при цьому одношарові клітини, з'єднані зі своїми сусідами десмосомами та щільними переходами, епітелій випирає всередину або назовні, якщо він не може рости ширше. Зовнішній ріст відомий як брунька, а внутрішнє зростання - це інвагінація. Руки і ноги ростуть з бруньок, тоді як зуби, волоски і мозок ростуть від інвагінацій. Це обидві форми інтерстиціального зростання.

морфогенез
Морфогенез - це поява нових форм в організмі. Це означає перейти від форми до якоїсь форми. Слово морфологія може означати вивчення цих форм або використовуватися як прикметник, що означає форму чогось (Наприклад, розщеплення піднебіння - це шанс на здорову морфологію піднебіння.) Від зиготи до стадії бластули (іноді його називають зародковими стадіями) куля клітин аморфний, він не має форми (крім сфери). Однак на початку гаструляції ембріон зазнає численних морфологічних змін. Нові форми виникають у ембріона, що розвивається. Розвивається всередині і зовні, хвостовий кінець і головний кінець, незабаром розвивається лівий і правий, і передній і задній. Після цього стає видно нервова система і кровоносна система. Як ви думаєте, скільки часу потрібно, перш ніж розвиватися зуби? Відповідь може вас здивувати.
 Малюнок 6.15: Д'Арсі Вентворт Томпсон запропонував, що те, що може виглядати як різні форми у споріднених видах, може бути перетворенням однієї основної картини. Кредит зображення: «Рис 150-153» Д'Арсі Вентворт Томпсон, Про зростання та форму знаходиться у суспільному надбанні, CC0
Малюнок 6.15: Д'Арсі Вентворт Томпсон запропонував, що те, що може виглядати як різні форми у споріднених видах, може бути перетворенням однієї основної картини. Кредит зображення: «Рис 150-153» Д'Арсі Вентворт Томпсон, Про зростання та форму знаходиться у суспільному надбанні, CC0Рис. 6.14 - знаменитий малюнок крихітних морських істот з мінералізованими екзоскелетами. Пізніше цей образ був проаналізований математичним біологом на ім'я Д'Арсі Вентворт Томпсон, який помітив, що різні біологічні форми можуть бути різними перетвореннями однієї і тієї ж форми. Він запропонував, перетворення відбулися різними локалізованими темпами зростання - те, що підтримує наші сучасні знання про морфогенну сигналізацію - як він проілюстрував на рис. 6.15. Його розглядають як першу особу, яка описує морфогенез. Коли Томпсон подивився на крихітних морських істот, він не бачив різних форм, він побачив зміни в одній формі. Ці зміни відбулися протягом еволюційного часу і призвели до різних морських істот (які поділилися єдиним родом). У цій книзі ми розглядаємо різні частини тіла, які мають одне і те ж походження, але змінюються з часом розвитку, такі як волосяні фолікули та зубні нирки. Ідеї Томпсона були розширені математиком Аланом Тьюрингом, який придумав термін морфогенез. Інші його досягнення включають винахід інформатики та порушення нацистського коду під час Другої світової війни (за допомогою багатьох жінок-математиків у парку Блетчлі). На жаль, він покінчив життя самогубством після арешту, засудження та ув'язнення за гомосексуалізм британським урядом, який він допоміг врятувати. Погляд їхнього уряду на гомосексуалізм розвинувся з тих пір, ми можемо краще зрозуміти поточний погляд, слідуючи його родоводу.
Морфогенез і генетика
Щоб мати глибоке розуміння того, як працює морфогенна сигналізація, це допомагає зрозуміти деякі поняття генетики. Коли ми говоримо зрозуміти, ми не маємо на увазі запам'ятовувати конкретні деталі, зосереджуватися на поняттях. По-перше, зверніть увагу на те, коли ми обговорюємо морфоген, ми, ймовірно, маємо на увазі саму сигнальну молекулу. Іноді ми обговорюємо морфогенний сигнал, і цей додатковий сигнал слова має велику різницю. Мутації, що змінюють сигнали морфогену, включають мутації гена для самої молекули позаклітинного морфогену (якщо це білок, такий як FGF та BMP) або ферменти, які синтезують морфоген (як у випадку з ретиноєвою кислотою), рецептори поверхні клітин для морфогенів, цитоплазматичні секундні месенджери та фактори ядерної транскрипції. Обговорення рівнів сигналізації морфогенів позбавляє нас від складності включення всіх можливих цілей. Далі деякі мутації можуть бути посилення функції, наприклад, мутація до рецептора, який запобігає його відключення, і вони часто успадковуються в аутосомно-домінантному порядку (для створення сигналу, де він не повинен бути). Крім того, мутації втрати функції перешкоджають нормальному функціонуванню білків і часто успадковуються за аутосомно-рецесивним малюнком (потрібно, щоб обидві копії були несправними, щоб призвести до втрати сигналу там, де він повинен бути, достатньо одного хорошого алелю). Деякі мутації повністю припиняють або повністю запускають морфогенні сигнали. і в таких випадках ми б сказали, що тези мутації мають повне проникнення. Інші мутації можуть частково блокувати або частково почати морфогенні сигнали, і мати часткове або неповне проникнення, вираженість симптомів падає на спектр.

Дозрівання
Дозрівання - це процес, за допомогою якого нові форми, створені морфогенезом, продовжують розвиватися і набувати більш зрілу форму. До кінця 9 тижня сформувалися всі основні форми органів, і ми говоримо, що ембріональний період закінчився і починаються етапи плода. Плід піддається дозріванню, а дозрівання включає шматочки попередніх процесів (ще один спосіб сказати, що дозрівання повторює інші процеси розвитку).
Фьюжн
Іноді два окремих шматочка однотипної тканини стикаються один з одним і зливаються в єдину тканину. Для епітелію це вимагає, щоб клітини поділяли той же тип трансмембранного білка, відомого як молекула клітинної адгезії. Для сполучної тканини це вимагає правильного зв'язування інтегрину з фібронектином. На рис. 147 основна картина зеленого буття зовні і червоного в середині спочатку порушується злиттям двох структур. Існує три способи видалення зелених клітин з середньої області. Перший спосіб полягає в тому, щоб клітини отримували сигнали керівництва і змушували клітини рухатися. Деякі вказівки є відразливими, наприклад, відсутність правильних контактів між клітиною та клітиною або високі концентрації відштовхуючого морфогену. Інші наведені сигнали привабливі, такі як правильний контакт клітини з клітиною або позитивний морфоген керівництва. Чи є зв'язок клітина-клітина або морфоген привабливим або відразливим, залежить від того, як була запрограмована клітина-мішень (які рецепторні гени вона експресує). Клітини в неправильному розташуванні також можуть бути спонукані піддаватися апоптозу (див. Нижче). Нарешті, ще одним способом реорганізації клітин є спонукання їх до дедиференціації (повернення до стовбурової клітини) та диференціювання на правильний тип клітин. Це називається переходом; як ранній розвиток, так і розвиток зуба включають приклади епітеліал-мезенхімальних переходів і мезенхімальних до епітеліальних переходів.

Апоптоз
При будівництві високих будівель або великих кораблів будівельні ліси будуються спочатку, щоб дозволити будівельникам дістатися туди, де їм потрібно, а потім видаляють після завершення будівництва. Так само під час розвитку апоптоз є невід'ємною частиною побудови частин тіла. Багато тканин виростають більше клітин, ніж потрібно в зрілому вигляді. Щоб пройти дозрівання, непотрібні клітини піддаються апоптозу. Наприклад, під час росту кісткової тканини додаткова хрящова тканина виступає в якості каркаса для остеобластів. Показано на рис. 6.18, руки і ноги починаються як весла, і тільки при апоптозі в чергуються областях відбувається морфогенез пальців рук і ніг. Без достатнього апоптозу люди народжуються з перетинчастими пальцями рук або ніг, або страждають анкілоглосією (їх язик жорстко закріплений на підлозі рота вираженою мовною вуздечкою).
ранній розвиток

Передімплантаційний період
Передімплантаційний період починається з запліднення, охоплює тиждень або близько того розвитку і закінчується імплантацією ембріона в матку. Запліднення - це об'єднання одного сперматозоїда і однієї яйцеклітини - потрібні тисячі сперматозоїдів, але в яйцеклітину допускається лише один. Цей союз з'єднує 23 материнські хромосоми з 23 відповідними батьківськими хромосомами, утворюючи зиготу. Запліднення найчастіше відбувається у напрямку до дистального кінця маткової труби. Протягом наступного тижня зигота піддається мітозу і збільшується кількість клітин, але не збільшується в розмірах. Всі амінокислоти та нуклеотиди для створення нових клітин протягом цього першого тижня надходять з цитоплазми зиготи, саме тому яйцеклітина є дуже великою клітиною. Після імплантації матка забезпечує сировину для того, щоб ембріон збільшився.
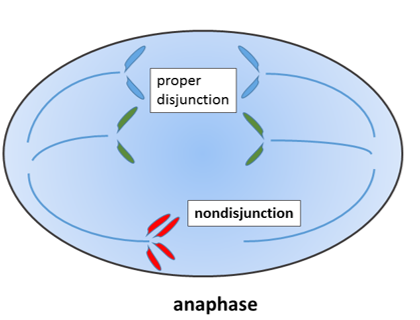
Під час мітозу 23 пари хромосом дублюються і поділяються між 2 дочірніми клітинами рівномірно. Якщо допущені помилки і непарна кількість хромосом потрапляє в дочірню клітину, це не дає можливості контрольної точки клітинного циклу і викликає апоптоз. Якщо хромосома зламається, це може привести до такого ж результату.

Однак є винятки. Клітина з непарним числом хромосоми 21 або 23 може вижити. Якщо ця клітина є яйцеклітиною, то зигота і всі наступні клітини матимуть непарну кількість хромосом. Це призводить до вроджених порушень, таких як трисомія 21. Якщо помилка хромосомної сегрегації трапляється на 2-клітинній стадії розвитку, то тільки половина клітин у дорослої людини мала б зайву хромосому, і стан було б менш важким. Зайві хромосоми перешкоджають процесам прийняття рішень в розвитку. Люди з трисомією 21 часто розвивають мікродонтію і можуть страждати від гіпослиновиділення та бруксизму. Тому вони потребують додаткового догляду, підтримуючи здорову гігієну порожнини рота.

Ранні поділи
Протягом першого тижня розвитку зигота проходить 3 відділи розщеплення, де мітоз відбувається синхронно (дивіться круте відео тут). Клітини дублюються по одній площині, 3 рази, виробляючи 8-клітинний ембріон. Синхронізація мітозу вимагає зв'язку між клітиною та клітиною між розривними переходами для координації контрольних точок клітинного циклу. Синхронізований мітоз призводить до того, що ембріон розвивається рівномірно. Той факт, що кілька клітин діляться в одному напрямку, означає, що ці клітини мають деяку полярність, навіть якщо ембріон в цілому ще не має видимого верху або низу. У міру продовження мітозу кількість клітин продовжує подвоюватися, але клітини не діляться по одній площині. Цей твердий куля клітин називається морулою. Оскільки мітоз триває, кількість клітин подвоюється, а клітини продовжують ставати меншими, поки тверда кулька клітин не стане порожнистою кулькою клітин, названих бластулою. Від зиготи до стадії бластули жодна з клітин не виявляється різною, крім як за розміром. Більш химерний спосіб сказати це те, що немає ніяких змін в морфології. Під час стадії бластули група клітин, названих ембріональним диском, відходить від інших. Доля ембріонального диска полягає в тому, щоб стати ембріоном, тоді як зовнішнім клітинам, відомим як трофобласт, судилося стати екстраембріональними структурами, такими як плацента. Бластула імплантується в ендометрій приблизно через тиждень після запліднення і продовжує розвиватися.

Гаструляція
Це не народження, шлюб або смерть, а гаструляція, яка воістину є найважливішим часом у вашому житті. —Льюїс Вольперт.
Огляд гаструляції
Під час гаструляції деякі клітини інвагінують, утворюючи шар клітин зсередини і шар клітин на зовнішній стороні ембріона. Значення цього полягає в тому, що це перший випадок, коли ембріональні клітини відрізняються один від одного. Але, почекайте, якщо всі клітини бластули ідентичні, чому мігрували лише деякі з них? Мітоз дійсно генерує 2 клітини з ідентичною ДНК, але не обов'язково ідентичною цитоплазмою. Якщо яйцеклітина має мРНК для фактора транскрипції, згрупованого в одну пляму цитоплазми, перше поділ розщеплення виробляє дві неідентичні клітини, одна з фактором транскрипції мРНК і одна без. Пізніше, коли мРНК перекладається, це може спонукати цю клітину мігрувати всередину, тоді як інші клітини залишаються зовні. Це материнський ефект, оскільки мРНК для фактора транскрипції не транскрибується з ДНК зигот, а виробляється клітинами яєчників мами і відкладається в яйцеклітину. Повернемося до гаструляції: кінцевим результатом гаструляції є ембріон з двома шарами (зовні і всередині). Уявіть собі, що тримаєте тенісний м'яч і штовхаєте великі пальці всередину, змушуючи частину зовнішньої гуми складатися всередину. Трубка, яка утворюється, стає шлунково-кишковим трактом, а отвір (можна здогадатися?) стає заднім проходом. Тому ембріон тепер має чіткий напрямок хвоста та напрямок голови. Рис. 6.23 представляє спосіб гаструляції морського їжака. Людські ембріони виглядають інакше, оскільки гаструляції піддається лише ембріональний диск, а не вся бластула. Але 2 шари однакові, людські гаструли просто виглядають більш розчавленими. О, і морські їжаки не утворюють анус-перший. Причина гаструляції морського їжака виглядає простішою, ніж гаструляція людини, полягає в тому, що бластули людини резервують деякі клітини (трофобласт), щоб сформувати плаценту та амніон, тоді як морські їжаки скидають свої яйцеклітини в море. Ми жертвуємо невеликою точністю тут, щоб зосередитися на важливих концепціях, поділених між видами. Якщо вам цікаво, дивитися фільми про ці процеси у людини не представляється можливим, але це для таких організмів, як ксенпозні лаевіси (жаби).
Далі клітини з зовнішнього шару мігрують між двома шарами. Це дає ембріону 3 шари, які є 3 зародковими зародковими шарами (рис.6.23). Зовнішній шар ектодерми і внутрішній шар ентодерми залишаються епітеліальними на вигляд, в той час як середній шар клітин (мезодерма) зазнає морфологічної зміни, і стає мезенхімою. Ця міграція починається в одному місці, званому вузлом Хенсена, який рухається вниз ембріона, утворюючи примітивну смугу позаду нього (рис. 6.24). Стороні ембріона з вузлом судилося стати спинною стороною, а смуга ділить ембріон на ліву половину і праву половину. Ви тепер стали свідками, нехай і в мультяшній формі, як щось (всі основні напрямки тіла) виникло з нічого. Не дозволяйте значенню цього прокрастися повз непоміченим. Вченим важко пояснити, що було до того, як утворився Всесвіт, або як Земля перейшла від життя до життя. Ембріологи, навпаки, пояснюють, як ембріони змінюються від відсутності форми до форми дуже детально (ми мали перевагу безпосередньо спостерігати та маніпулювати цим процесом).
| 3 ембріональні шари | осередок долі |
|---|---|
| Ектодерм | Епітелій шкіри та слизової оболонки порожнини рота, нервової тканини |
| Мезодерма | Сполучна і м'язова тканина |
| Ендодерм | Епітеліальна вистилання порожнистих органів |
Таблиця 6.2: Три зародкові шари розвиваються під час гаструляції

Епітеліал-мезенхімальний перехід
Процес, за допомогою якого ектодермальні клітини перестають бути ектодермою і мігрують в середину і утворюють мезодерму, називається епітеліал-мезенхімальним переходом (ЕМТ). Епітеліальні клітини ектодерми повинні спочатку втратити свої клітинно-клітинні контакти з сусідніми клітинами, що робить їх менш епітеліальними. Вони втрачають полярність, дедиференціюються в стовбурову клітину, потім диференціюються в мезенхімальні стовбурові клітини. Цей процес потрібен не тільки для формування мезодерми під час гаструляції, він відбувається під час міграції нервового гребеня, загоєння ран та метастазування раку. Клітини мезодерми (назва середнього шару) утворюють мезенхіму (тип тканини), тоді як інші два шари залишаються епітеліальними. Існує зворотний процес, званий мезенхімальним переходом в епітеліальний (МЕТ), який також відбувається під час загоєння ран. ЕМТ дозволяє епітеліальним стовбуровим клітинам зі здорової ділянки слизової оболонки ротової порожнини перетворюватися в мезенхімальні стовбурові клітини, мігрувати від здорової області в пошкоджену область. MET дозволяє мезенхімальним стовбуровим клітинам перетворюватися назад в епітеліальні стовбурові клітини, які діляться і диференціюються на нові кератиноцити. Оскільки це імітує те, що відбувається під час гаструляції, ми говоримо «загоєння ран повторює (знову станує) розвиток».
Диференціація ектодерми

Нейруляція
Гаструляція дала ембріону його першу полярність, а також початок шлунково-кишкового тракту. Однією з наступних систем органів для розвитку є центральна нервова система. Основні процеси, що використовуються при нейруляції, повторно використовуються при формуванні інших порожнистих органів. По-перше, що лежить в основі мезодерми, відома як нотохорда, посилає морфогени сусіднім ектодермальним клітинам. Це спонукає прилеглу область ектодерми диференціюватися на нейро-ектодерму з подальшою місцевою проліферацією. Оскільки ці епітеліальні клітини не можуть легко поширюватися з боку в бік, вони інвагінують. Зрештою, інвагінаційні нейро-ектодермальні клітини втрачають свої контакти з ектодермою і зливаються з іншими нейро-ектодермальними клітинами, створюючи нову структуру, звану нервовою трубкою. Ця трубка розвивається в головний мозок і хребет. Якщо вам цікаво про нотохорду, її клітинна доля полягає в тому, щоб в основному пройти апоптоз, хоча деякі залишаються як пульпоз ядра хребетних дисків. У крихітних риб'ячих хордових без хребта, званого амфіоксисом, нотохорда зберігається. Це буде не останній раз, коли ви побачите, як людський ембріон створює структуру, яка повністю відходить або змінюється.
Нервова трубка починає згортатися від передньої частини ембріона, і застібається в передньо-задньому напрямку. Блискавка не ідеальна, передній кінець також зростає ширше, оскільки він складається, утворюючи примітивний мозок. Для всіх процесів розвитку, які ми робимо і не охоплюємо, передній кінець, як правило, є областю, яка розвивається спочатку, а потім більше задніх областей. Застібка нервової трубки вимагає достатнього рівня фолієвої кислоти, тому важливо, щоб меланін матері захистив цей вітамін, інакше може призвести до неповного закриття нервової трубки (spina bifida).

Деякі з нейро-ектодермічних клітин не зливаються. Натомість ці клітини проходять епітеліал-мезенхімальний перехід і мігрують від нервової трубки. Ці клітини - це клітини нервового гребеня, їм судилося стати низкою важливих клітин і тканин по всьому організму, включаючи меланоцити, одонтобласти, зубну пульпу, цементобласти та нейро-мезенхіму дуг глотки. Ці клітини іноді називають четвертою ембріональною тканиною (на додаток до трьох, що виникають під час гаструляції), що говорить про те, що деякі люди вважають, що нервовий гребінь важливий. Навіть після того, як клітини нервового гребеня мігрують до нових і віддалених тканин, вони часто зберігають видимі ознаки своєї нервової лінії, такі як дендрити на меланоцитах або одонтобластний процес одонтобластів. Щоб полегшити міграцію, клітини нервового гребеня експресують фермент, який називається матричною металопротеїназою (MMP). Цей фермент перетравлює білки, знайдені в позаклітинному матриксі. До теперішнього часу ми сподіваємося, що згадати фразу загоєння ран повторює розвиток, ферменти матриксу металопротеїнази повторно використовуються для загоєння певного типу зубних ран. Коли клітини нервового гребеня досягають глоткових дуг, вони виділяють морфогени FGF і BMP, які антагонізують один одного, виробляючи смугастий малюнок уздовж ектодерми. Деякі регіони спонукають до утворення зубних бруньок, між ними утворюються слизові оболонки порожнини рота.
Диференціація мезодерми
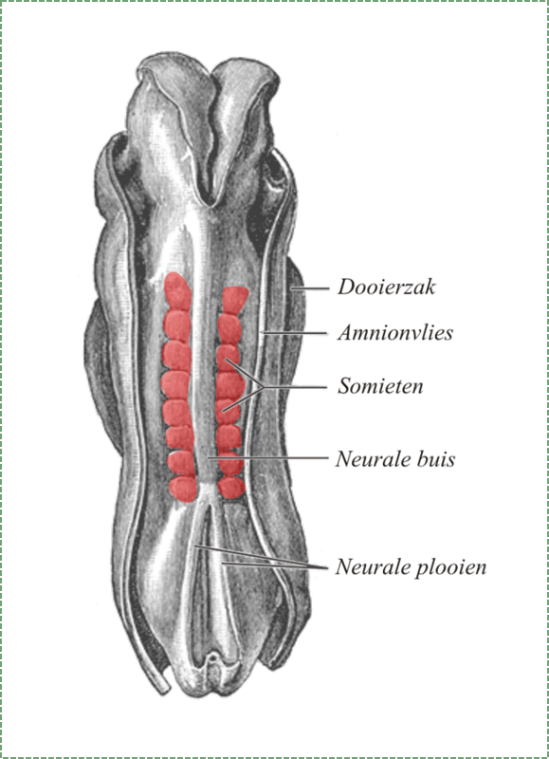
утворення сомітів
Протягом того ж часу, що і нейруляція, мезодерма піддається диференціації. Мезодерма починається як аморфний шар мезенхіми. Потім ділянки мезенхіми відщипують у повторюваних сферичних структурах, званих сомітами, утворюючи сегменти вздовж передньо-задньої осі ембріона. Доля сомітів полягає в тому, щоб стати твердими органами, або повторюючи одиниці сполучної тканини, такі як ребра та хребці, або повторюючи одиниці м'язової тканини, такі як пряма черевна порожнина та міжреберні м'язи. Формування кожного соміту передбачає мезенхімальний перехід до епітелію, деякі мезенхімальні стовбурові клітини диференціюються в епітелій, який відокремлює один соміт від наступного.

формування серця
На третьому тижні мезодерма також починає розвиватися в кров, серце і кровоносну систему. Мезенхімальні стовбурові клітини спочатку диференціюються на острівці крові. Далі починається ангіогенез. Два великих кровоносних судини fuse— з деякою допомогою клітин нервового гребеня - утворюють примітивне серце, яке починає битися.
Диференціація ентодерми
формування глотки
Ентодерма, яка мігрує внутрішньо під час гаструляції, розширює свій шлях до переднього кінця ембріона. При цьому утворюється примітивний передок, який згодом стає глоткою, ротовою порожниною і порожниною носа. В цей час, однак, немає отворів для рота і ніздрів.

4-й тиждень
Ми зараз тільки до 4-го тижня розвитку. Багато що сталося! У цей час ембріон продовжує зазнавати локальних областей проліферації, малюнка, злиття та інших процесів, охоплених на початку глави. Це призводить до утворення рудиментарних (основних) структур. Коли структура вперше стає видимою з поверхні ембріона, її часто називають плакодом (ектодермальне потовщення). Клітинам в плакоді судилося стати органом, але їх загальна форма - не більше, ніж піднята шишка або неглибока яма. До 4-го тижня починають розвиватися пов'язані з головою органи, перераховані в таблиці 6.3.
| Структура | Доля |
|---|---|
| Плакоди очей | примітивні очі |
| Otic плакати | примітивні вуха |
| Носові плакати | примітивний ніс |
| дуги глотки | так багато речей вони отримують власну главу |
| Ротоглоткова мембрана | примітивний рот (ще не відкритий) |
Таблиця 6.3: примітивні структури, сформовані протягом 4-го тижня розвитку
Клінічні застосування раннього розвитку
Порушення на цих ранніх стадіях розвитку зазвичай настільки важкі, що ембріон не виживає. Однак існують клінічні умови, пов'язані з цими ранніми стадіями та здоров'ям порожнини рота.
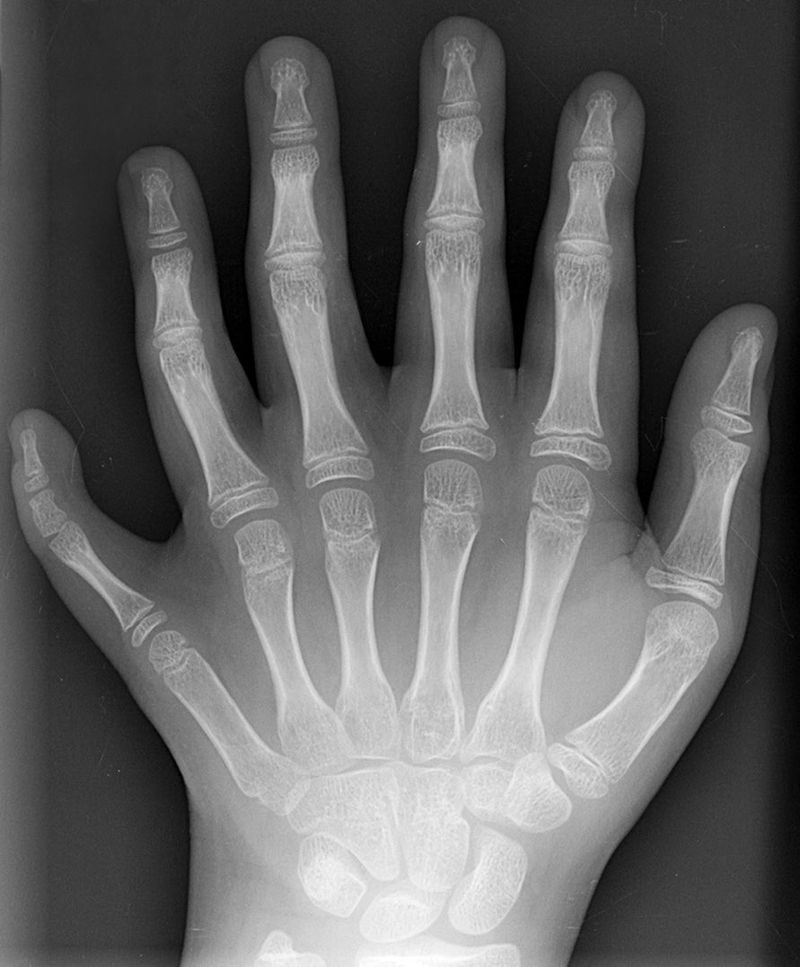
Мутації генів гомеобox
Мутації гена гомеобкса можуть призвести до того, що цілі структури відсутні або розвиваються в неправильному місці. Ранні експерименти на мухах привели до спостережень за ногами, де повинні розташовуватися вусики, або вусиків, де повинні розташовуватися крила. Існує кілька генетичних порушень, пов'язаних з гомеобаком, у людини. Порушення, пов'язані з гомеобіксом, рідкісні, оскільки мутації, як правило, смертельні, але коли вони не є, умови часто важкі. Невеликий список можна знайти в таблиці 6.4
| Синдром | Задіяний ген гомеобox | Симптоми включають |
|---|---|---|
| Синдром Аксенфельда-Рігера | ЛИСИЦЯ 1, ПІТХ2 | Переважно пов'язані з очима, іноді гіподонтія та мікродонтія |
| Аутосомно-домінантна синдактилія | СКРИНЬКА 13 | Синдактилія (зрощені пальці рук або ніг) |
| синдром Гуттмахера | ХОКСА13 | Полідактилія (додаткові пальці рук або ніг) |
| Ектодермальна дисплазія | HOXC13 | Дивіться нижче |
Таблиця 6.4: Кілька прикладів гомеобосних мутацій у людини.
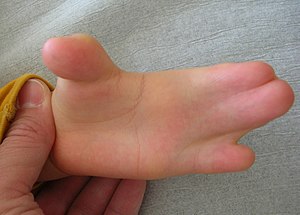
Синдром Аперта
Синдром Аперта викликаний мутацією в рецепторі морфогену FGF. Він класифікується за широким спектром симптомів, включаючи черепні деформації та синдактилію. ФГФ бере участь у формуванні апарату глоткової дуги (розглянуто в наступному розділі), що пояснює черепно-лицьові аномалії. Актуальною для цієї глави є здатність FGF (як і багато факторів росту) пригнічувати апоптоз. Наявність мутації, яка призводить до того, що рецептор FGF весь час пригнічує апоптоз у лопатках для рук та ніг. Області апоптозу зобов'язані виробляти пальці рук і ніг, отже, ця мутація призводить до синдактилії. Часткові порушення сигналів FGF і BMP можуть призвести до часткової синдактилії або лямки пальців рук або ніг.
 Малюнок 6.35: Малюнок 185: «Майкл Берріман, актор» Стефана Боргграфе ліцензується відповідно до CC BY 4.0
Малюнок 6.35: Малюнок 185: «Майкл Берріман, актор» Стефана Боргграфе ліцензується відповідно до CC BY 4.0Ектодермальна дисплазія (частина 1 з 2)
Проблеми з індукцією клітин нервового гребеня при нейруляції призводять до порушень в утворенні зубів, волосяних фолікулів, слинних залоз і інших структур. Спільним є те, що всі вони є спеціалізованими структурами ектодерми, індукованими клітинами нервового гребеня для диференціації. У здорового ембріона ектодермальні клітини отримують морфогени, які активують або інактивують правильні фактори транскрипції, щоб викликати диференціацію на клітини нервового гребеня. Нейронні клітини гребеня мігрують у віддалені області тіла, визначають їх розташування, взаємодіючи з морфогенами в наземній речовині, і вивільняють інші морфогени, щоб індукувати ділянки ектодерми диференціюватися на потові залози, слинні залози, зубні бруньки або волосяні фолікули. Мутації в будь-якому з генів цих морфогенів, їх рецепторних білків або факторів транскрипції нижче за течією можуть призвести до стану під назвою ектодермальна дисплазія. Ця група синдромів зустрічається рідко, лише 7000 випадків у всьому світі, але в цих синдромах задіяно щонайменше 40 різних генів. Порівняйте це з серповидно-клітинною хворобою, яка в даний час вражає понад 100 000 американців (переважно афроамериканців, латиноамериканців, греко-американців, турецьких американців та італійських американців), все через мутації в одному гені, гемоглобін-бета. Сенс цього порівняння полягає в тому, щоб виділити, коли ці гени виражені. Індукція ектодермальних стовбурових клітин для проліферації та диференціації на різні придатки є складною і відбувається під час ембріогенезу. Мутація в морфогенах, рецепторах морфогену, других месенджерах або факторах транскрипції та генах нижче за течією, які активуються для індукування диференціації, є можливими мішенями, які викликають ектодермальну дисплазію. Мутації цих генів у клітинці нервового гребеня призводять до порушення в будь-якій з наступних клітин, індукованих цим новим типом клітин, подібно до способу боротьби з носієм м'яча в футболі (футболі) порушує геймплей будь-якої з його потенційних мішеней проходження. Для порівняння, тільки еритроцити експресують гемоглобін-бета, і вони є невиліковно диференційованими клітинами, вони не стають будь-яким іншим типом клітин. Очевидно, еритроцити є важливим типом клітин, який можна знайти на початку ембріогенезу, але симптоми серповидноклітинної хвороби мають менший спектр, ніж типи захворювань, про які ми обговорювали, такі як ектодермальна дисплазія.
 Малюнок 6.36: Гейтен Джон Матараццо III, актор і активіст ПЗС. Зображення кредиту: Гейтен Джон Матараццо III, Гейдж Скідмор, має ліцензію CC BY SA 3.0
Малюнок 6.36: Гейтен Джон Матараццо III, актор і активіст ПЗС. Зображення кредиту: Гейтен Джон Матараццо III, Гейдж Скідмор, має ліцензію CC BY SA 3.0
Клейдо-черепний дизостоз
Клейдо-черепний дизостоз (ПЗС) - вроджене порушення, викликане мутацією до фактора транскрипції, необхідного для диференціації кістки та зубів. Потрібно викликати клітини остео-хондро-попередника для виходу з клітинного циклу і диференціювання в остеобласти. Він також повторно використовується для індукування диференціації одонтобластів. Крім того, після того, як зуби сформувалися, цей транскрипційний фактор повторно використовується для активації експресії ферменту Matrix металопротеїнази, який необхідний для ремоделювання альвеолярних розеток. Без цього ферменту відбувається ретенція молочних зубів. Зубні імплантати або зубні протези (такі як ті, які актор і CCD-філантроп Gaten John Matarazzo III отримали на рис. 6.36) є кращим лікуванням. Крім того, у людини з ПЗС можуть бути невеликі ключиці і зміни форми черепа — кісток, які утворюються шляхом внутрішньоперетинчастої окостеніння. Це ілюструє дві основні концепції в розвитку. Спочатку багато структур утворюються в одну сторону, але потім переробляються, щоб виконувати іншу функцію (зуби утворюються шляхом складання всередину, пізніше вони рухаються назовні). По-друге, багато різних закономірностей в ембріології повторно використовуються (рекапітулюються), такі як видалення тканини під час міграції нервового гребеня і видалення тканини під час прорізування зуба.
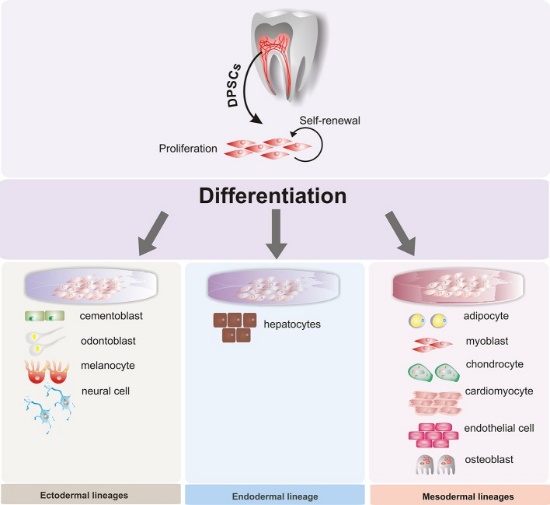
Терапія стовбуровими клітинами
Коли клітини невиліковно диференціюються, вони постійно інактивують непотрібні гени шляхом метилювання та зберігання навколо гістонів. Дослідники вивчають способи змінити цей процес та направляти диференційовані клітини, щоб повернутися до стану стовбурових клітин. Це підвищує можливість сприяння регенерації тканин, які інакше не регенеруються, усуваючи необхідність тканинної трансплантації або трансплантації. Оскільки деякі зубні тканини погано регенеруються, існує потенціал для застосування цих технологій до порожнини рота, наприклад, вирощування біологічних зубних імплантатів замість використання металів та кераміки. Однак більший інтерес був викликаний придбанням мезенхімальних стовбурових клітин із зубних тканин. Наприклад, стовбурові клітини, виділені з верхньощелепних третіх молярів, були використані в клінічних випробуваннях для поліпшення загоєння та зменшення потреби в трансплантованій тканині в щелепно-лицьовій хірургії. Через їх потенціал диференціюватися на широкий спектр клітин, великий інтерес був поставлений до збору зубних стовбурових клітин для лікування захворювань, не пов'язаних з порожниною рота. При правильному морфогені та великій кількості стовбурових клітин зубів можна змінити пошкодження, спричинені нейродегенеративними захворюваннями, такими як хвороба Альцгеймера (AD) та хвороба Паркінсона (PD), травма хребта, інфаркт міокарда (інфаркт) та м'язова дистрофія (MD). Лінія клітин, що виробляють дентин, пульпу, цемент і пародонт, допоможе пояснити зв'язок з нейродегенеративними розладами. Що ще ви будете робити з витягнутими третіми молярами? На жаль, в США немає юридичного визначення того, що являє собою безпечну, ефективну терапію стовбуровими клітинами. Навіть якщо будуть введені хороші правила, медико-туризм не дотримується. Окрім чесних та хороших досліджень у галузі терапії стовбуровими клітинами (Інститут стовбурових клітин та регенеративної медицини Вашингтонського університету є хорошим прикладом), існує велика кількість сумнівних клінік стовбурових клітин, що працюють на сірому ринку, використовуючи ризиковані, неперевірені методи і пред'являючи абсурдні претензії взяти якомога більше грошей у вразливих людей. Отже, хто є хто? Терапії, які використовують або підвищують власні стовбурові клітини пацієнта, мають найкращі шанси забезпечити позитивні результати з мінімальним ризиком побічних ефектів - особливо якщо ця терапія спрямована, наприклад, шляхом додавання певного морфогену для підвищення певного результату. З іншого боку, викидання екзогенних стовбурових клітин на проблемні клітини інших людей, інших тварин або з нетваринних джерел - несе найбільший ризик серйозних несприятливих наслідків з мінімальним потенціалом користі. Майте на увазі, що 19 з 20 (або близько того) потенційних наркотиків, які, як показано науково, працюють на тваринних моделям, зазнають невдачі в випробуваннях на людях. Це означає, що в публікації є наукові докази для лікування, які, як ми знаємо, не працюють, але інші люди можуть вибрати позитивну інформацію та використовувати її для продажу продукту.
Глава 5* Глава 7