1.1: Огляд клітинної біології
- Page ID
- 68203

- Огляд клітинної біології
- Усередині осередку:
- Клітинна мембрана, цитоплазма
- органели
- Поза осередку:
- Екстра-клітинна матриця (ECM)
- Екстра-клітинна рідина (ECF)
- Ні в, ні поза:
- Розподіл клітин
- загибель клітин
- З'єднання клітин

Огляд клітинної біології
Клітина - це основна одиниця життя. Це найменша річ, яку ми називаємо живими (не сперечаючись про віруси), а людський організм складається з 10 трильйонів з них. Ми починали як єдина клітина, і мета цього класу - дізнатися трохи про те, як ця клітина перетворилася на трильйонно-клітинний організм, яким ви є сьогодні. Майже всі інструкції для виготовлення трильйонів cells— як зробити їх, коли зробити їх, де зробити them— знаходяться в межах цієї однієї клітини. Різні клітини мають різні функції, хоча майже кожна клітина в одній людині має однакову ДНК. Щоб відрізнятися один від одного, клітини експресують різну ДНК. Це важливий процес, який називається диференціацією, який насправді не є більш складним у концепції, ніж клітини, що переходять від вигляду нудних родових клітин до пошуку та поведінки інакше, ніж інші клітини. Ми даємо різні назви клітинам, коли вони вносять ці зміни. Наприклад, стовбурова клітина, яка породжує, о, скажімо, смурф, буде названа стовбуровою клітиною Smurfal. Оскільки стовбурові клітини Смурфаля діляться та диференціюються на клітини, що утворюють смурф, клітини, які насправді роблять смурф, називатимуться Smurfoblasts, а коли Smurf буде закінчений, клітини всередині цього Смурфа будуть називатися Смурфоцитами. Стовбурові клітини, знайдені в тканині, званої мезенхімою, називаються мезенхімальними стовбуровими клітинами, клітини, що виробляють дентин, називаються одонтобластами, клітини в зрілому цементумі називаються цементоцитами. Зверніть увагу, що диференціація - це не один крок, а ряд кроків уздовж спектра, від найменш диференційованого, до більш диференційованого, поки ми не досягнемо невиліковно диференційованої клітини. Метою цієї глави є огляд аспектів класу клітинної біології, які з'являються в цьому підручнику, і не набагато більше. Ми припускаємо, що ви охопили клітинну біологію в обов'язковому порядку до цього класу. Нам потрібно переглянути частини клітини, які допомагають нам пояснити складні поняття, такі як диференціація та розвиток. Якщо ви виявите, що вам потрібно більше, ніж швидке оновлення, ось кілька посилань від NIH, які мають ряд дуже корисних публікацій та відео (безкоштовно), таких як:
- 3D анімації людської клітини
- Вивчіть генетику (інтерактивний сайт)
- Всередині комірки (завантаження pdf та epub)
Дві корисні (і безкоштовні) електронні книги, які ви можете завантажити, є з OpenStax:

Усередині клітини
Клітинна мембрана і цитоплазма
Кожна клітина людини оточена плазматичною мембраною. Плазмова мембрана відокремлює клітину від її середовища, і дозволяє певним матеріалам проникати і виходити з клітини. Фосфоліпіди і холестерин утворюють бар'єр, який відокремлює клітину від зовнішнього середовища, що, в свою чергу, дозволяє нам вказувати на цю клітину і називати її річчю. Транс-мембранні білки охоплюють плазматичну мембрану і регулюють те, що йде всередину або назовні. Інші трансмембранні білки отримують сигнали від інших клітин або навколишнього середовища, і передають цю інформацію всередину клітини. Треті опосередковують прикріплення до структур поза клітиною. Інші інтегральні мембранні білки розташовані на зовнішній або внутрішній поверхні плазматичної мембрани, але не охоплюють всю ширину фосфоліпідного бішару. Багато класів біології зосереджені в значній мірі на ядрі, тенденція, яку наполягав один з перших афроамериканців, який отримав ступінь доктора філософії, доктор Ернест Еверетт Джаст, у своїй книзі «Біологія поверхні клітин». Доктор Джаст був ембріологом, і в ембріології білки плазматичної мембрани, особливо рецептори і білки прикріплення - дуже важливі. Цитоплазма - це желатинове наповнення клітини, і іноді її називають внутрішньоклітинною рідиною (МКФ). Цитоплазма включає поживні речовини і електроліти, поглинені з рідини, що оточує всі клітини. Крім того, цитоплазма включає в себе ряд білків і глікопротеїнів, синтезованих клітиною. Ці молекули можуть мати і інші важливі функції, але вони притягують воду з ЕКФ. Кінцевим результатом є те, що клітини мають желатинову начинку, а не водянисту. Ця желатинова начинка наповнена рядом органел, подібно до того, як в салаті з желе моєї бабусі містив виноград, родзинки та інші харчові або харчові речовини, які жодним чином не перетворили це желе в справжній салат.
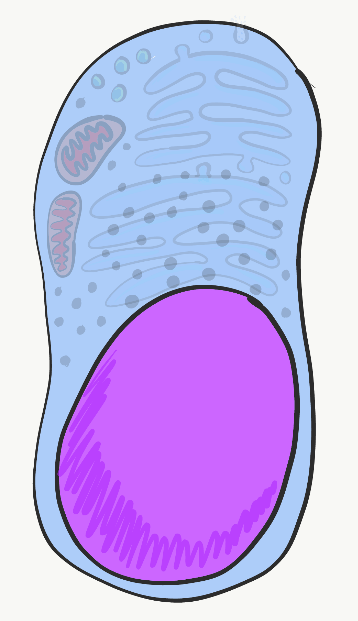
Ядро
Ядро містить практично всю дезоксирибонуклеїнову кислоту клітини (ДНК). ДНК - це інструкція з виготовлення копії рибонуклеїнової кислоти (РНК), і більшість РНК перекладається в цитоплазмі на білки, знайдені всередині і зовні клітини. Це також інструкція про те, коли і де робити ці білки. Наприклад, епітеліальні клітини слизової оболонки рота не виробляють ферменти, які виділяють кальцій і фосфат у позаклітинний матрикс. З іншого боку, епітеліальні клітини, які диференціюються на амелобласти, роблять ці білки, експресуючи ДНК для цих ферментів, після того, як їм сказали це зробити інші клітини, які називаються клітинами нервового гребеня. Всі клітини в організмі людини мають однакову ДНК (за кількома винятками). Однак різні клітини експресують різну ДНК в різний час. ДНК можна розділити на 2 основних типи. Є гени, кожен ген більш-менш інструкція для окремого білка. Решта ДНК складається в унікальні форми, які дають вказівки щодо того, коли і де експресувати ці гени. Останню відносять до некодуючих ДНК. Для експресії ДНК білки, звані факторами транскрипції (за допомогою спеціальних малих молекул), зв'язуються з регуляторними областями ДНК, відкривають відповідну область та набирають ферменти, які копіюють одну нитку ДНК в месенджерну РНК (мРНК), яка залишає ядро бути перекладається на білок. Інші фактори транскрипції можуть тимчасово пригнічувати експресію генів.
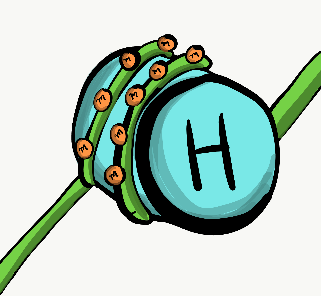
Фактори транскрипції вмикаються і вимикають гени швидко, у відповідь на зміни в середовищі клітини. Але коли клітини диференціюються, вони закривають гени, які їм не потрібні більш постійно. Замість того, щоб покладатися на включення та вимикання факторів транскрипції, що зв'язуються з регуляторними областями ДНК, ці інактивовані гени метилюються (приєднується група -CH 3), упаковуються навколо гістонових білків і поміщаються в тривале зберігання. Під час мітозу копіюється картина метилювання ДНК та упаковки гістону. Це означає, що модель генів, які доступні або упаковані далеко, успадковується обома дочірніми клітинами, але оскільки це успадкування не є різницею в послідовності ДНК, воно відоме як епігенетичне успадкування. Епігенетичні риси більш здатні модифікуватися у відповідь на зміни навколишнього середовища, ніж гени (ДНК), і відіграють важливу роль у диференціації клітин та долі клітин, що більш детально розглянуто в розділах 8-11. У нас в ядрі 46 молекул ДНК - це химерно довгі молекули, але лише 46 у кількості - 23 материнські та 23 батьківські. Під час мітозу ці 46 молекул щільно упаковані в 46 хромосом (раз 2). Ця упаковка передбачає використання гістонів майже по всій довжині ланцюга ДНК і дозволяє хромосоми бачити під світловим мікроскопом. В інший час ДНК в основному розмотується (непотрібні інструкції намотуються навколо гістонів, решта вільні для транскрибування), і заповнює ядро таким чином, що не виглядає дуже захоплюючим. Ми називаємо це хроматином ДНК, і функціонально він набагато більш захоплюючий з двох форм.

Рибосоми
Видимі по всій цитоплазмі невеликі цятки, виготовлені з білка і РНК, звані рибосомами. Ці структури переводять інструкції мРНК, що надійшли з ядра, а це означає, що інструкції з мРНК використовуються для керівництва зв'язком ряду амінокислот у довгий білок. Групи з трьох нуклеотидів мРНК, які називаються кодонами, вказують рибосомі, яку амінокислоту додати до білка. Як і регуляторні області ДНК, РНК, виявлена в самій рибосомі - не інструкції з мРНК, а рибосомної рРНК, є типом РНК, який складається в специфічні форми, які разом з рибосомними білками виконують хімічну реакцію перекладу. Вільно плаваючі рибосоми в цитоплазмі синтезують білки, які залишаються в цитоплазмі, такі як кератин або ферменти, які опосередковують апоптоз.
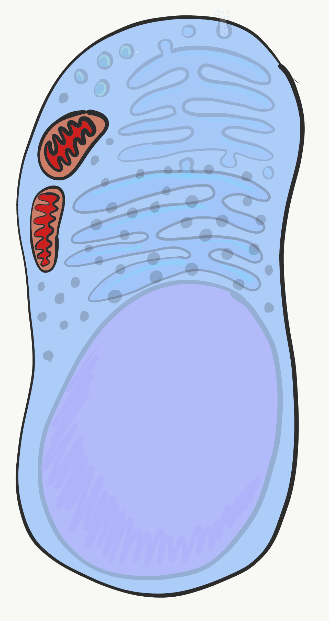
Мітохондрії
Мітохондрії - це місце, де виробляється більшість аденозинтрифосфату (АТФ). АТФ виготовляється з аденозину, плюс три фосфатні (PO 4 3-) групи - зверніть увагу на фосфатну частину, він також є основним компонентом кісток, емалі, дентину та цементу. АТФ живить практично всі клітинні процеси, включаючи транскрипцію і трансляцію слизових білків всередині слинної залози, електричні сигнали, що посилаються нейронами на мові при попаданні їжі в ротову порожнину, і скорочення міо-епітеліальних клітин, щоб викликати слиновиділення. Мітохондрії спалюють глюкозу, використовуючи кисень, і запрягають частину енергії, що виділяється у вигляді АТФ. Мітохондрії відрізняються від інших органел тим, що містять трохи власної ДНК, яка успадковується якраз від матері. Мітохондрії також містять дві фосфоліпідні бішарові мембрани, не одну, як інші органели. Вони використовують цю додаткову мембрану для генерації АТФ. Можливо, ви раніше охоплювали гліколіз і цикл лимонної кислоти. Частина, яку слід пам'ятати, полягає в тому, що мітохондрії використовують протонний (або H +) градієнт, який робить внутрішню частину мітохондрій кислою, а отже, потенційно токсичною для решти клітини.
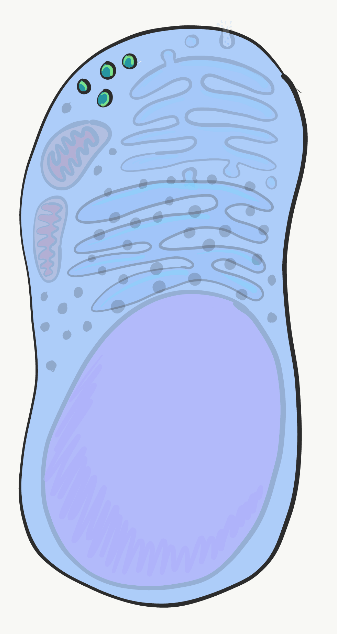
Лізосоми
Лізосоми - це невеликі відсіки, оточені тими ж фосфоліпідами, що і в плазматичній мембрані. Усередині лізосом знаходяться кислоти та травні ферменти, які можуть бути використані для знищення речей всередині клітини, коли вона зношується, або матеріали, які клітина з'їла ззовні (наприклад, сміття, бактерії). Коли клітина гине і починає розпадатися, сусідні клітини загрожують пошкодженням кислот і ферментів, що знаходяться в лізосомах. У ротовій порожнині епітеліальні клітини мають тривалість життя лише днів, перш ніж вони зношуються, тому цим клітинам дуже важливо спочатку нейтралізувати кислоти та ферменти у своїх лізосомах, у процесі, який називається апоптозом.

Ендоплазматичний ретикулум
Ендоплазматичний ретикулум (ER) - це серія з'єднаних між собою трубок, оточених фосфоліпідним бішаром - подібним до лізосом, тільки більшим, більш трубчастим і не наповненим кислотою. Гладкий ендоплазматичний ретикулум (SeR) - це місце, де клітини виробляють ліпіди та зберігають кальцій. Грубий ендоплазматичний ретикулум (ReR) покритий рибосомами. Білки, виготовлені цими рибосомами, потрапляють всередину ReR, потім подорожують до апарату Гольджі і або виділяються (наприклад, слизові білки) або залишаються в плазматичній мембрані (наприклад, білки-перехідні клітини або рецептори для молекул морфогену).

Апарат Гольджі
Апарат Гольджі - це ще один набір трубок, схожий на ReR. Невеликі мембранно-замкнуті сфери, звані бульбашками, човником білків, зроблених в ReR, до апарату Гольджі, де білки модифікуються. Часто ці білки мають цукру, прикріплені до них, що робить їх глікопротеїнами. Бульбашки забирають ці білки в плазматичну мембрану, де вони або секретуються, або стають частиною плазматичної мембрани. Ми покриваємо роль, яку виділяється білок колаген відіграє в емалі та пародонтальної зв'язці. Ми також охоплюємо спільну роль секретованого глікопротеїну фібронектину та зв'язаного з мембраною білкового інтегрину у загоєнні пошкодженої тканини ясен.

Цитоскелет
Цитоскелет - це мережа структурних білків всередині цитоплазми. Ця мережа надає клітині її форму, можливість змінювати свою форму, або мігрувати. Показано на рис. 1.10, клітини мають мікротрубочки і актинові нитки, забарвлені відповідно в червоний і зелений кольори. Ці білки, як правило, не видно на більш старомодних гістологічних images— обидва були б рожевими на плямі H&E, як і розчинні білки в цитоплазмі.
Поза осередку
| Позаклітинний матрикс: |
|---|
| Мелена речовина |
| Волокна |
Таблиця 1.1: Дві основні компоненти ECM. Екстра-клітинна матриця (ECM) включає в себе весь матеріал, знайдений поза клітинами. Зазвичай він розбитий на два компоненти, перераховані в таблиці 1.1.
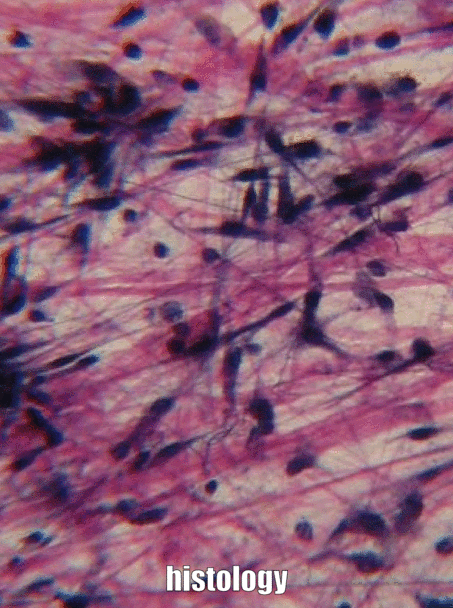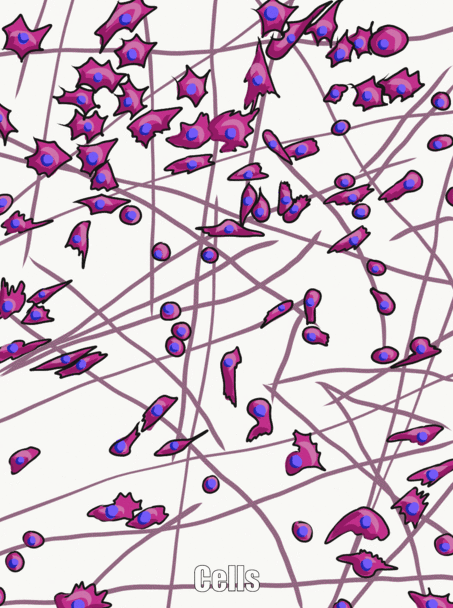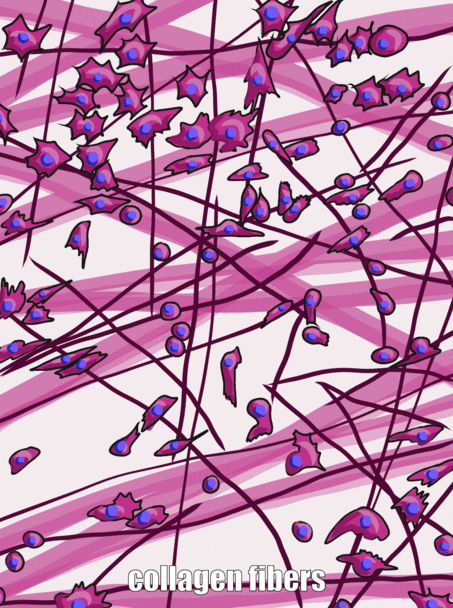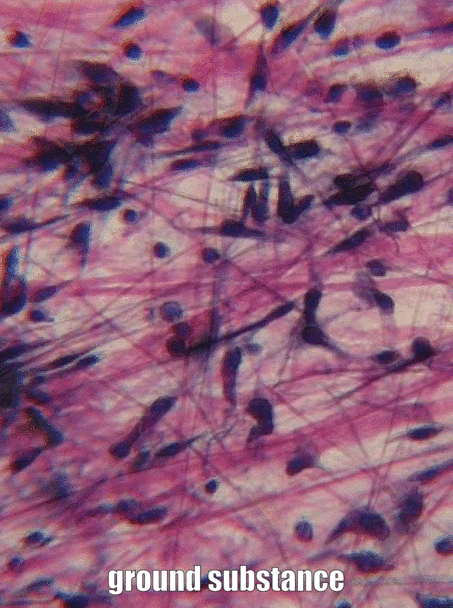
Мелена речовина
Мелена речовина - це назва для всього матеріалу поза клітиною, який не має особливої форми при розгляді під традиційною мікроскопією. Однією з частин наземної речовини є екстра-клітинна рідина (ECF), яка є водою та поживними речовинами, оточуючими клітинами, які називали плазмою, коли вона перебувала всередині кровоносної судини. Як тільки рідина виходить з крові і оточує клітини, вона називається ECF. На відміну від плазми, ця рідина утримується на місці іншою основною частиною подрібненої речовини, до складу якої входять білки, глікопротеїни та полісахариди. Ці молекули є розчиненими речовинами, а їх розмір робить їх нерухомими, тому вони добре утримують рідину на місці, утворюючи гель. Це не схоже на багато під мікроскопом, не більше, ніж якби ви подивилися дуже уважно на якесь желе. Ці білки, глікопротеїни і полісахариди виробляються і секретуються клітинами (ці клітини, ймовірно, мають багато апаратів ReR і Гольджі). Одним з глікопротеїнів, що містяться в подрібненій речовині, є фібронектин, який є дійсно довгим білком. Клітина може розпізнавати, зв'язуватися з фібронектином і рухатися вздовж, якщо вони мають правильний білок інтегран, що охоплює їх плазматичну мембрану. Тому фібронектин діє не тільки як дорога, по якій рухаються клітини, це ще й дорожня карта. Отримання потрібних клітин в потрібне місце в потрібний час дуже важливо як в оздоровленні, так і в розвитку. Насправді те, що ви дізнаєтеся про розвиток, повторно використовується (рекапітулюється) у зціленні. Ще однією важливою молекулою, що міститься в подрібненій речовині, є великий полісахарид під назвою гіалуронова кислота (HA). Як і фібронектин, клітини можуть зв'язуватися і подорожувати по ГК (використовуючи інший тип білка плазматичної мембрани), який має застосування в стоматології, наприклад, допомагаючи клітинам ясен прилипати до зубного імплантату і утворювати стійке до бактерій ущільнення. Ми не можемо побачити фібронектин або ГК без використання деяких сучасних прийомів візуалізації, тому вони перераховані як подрібнена речовина, а не в наступному розділі, волокна. Оскільки клітини мігрують над білками подрібненої речовини, ми говоримо, що білки ECM функціонують як ешафот. Без риштування тканини ростуть тільки з їх країв. Це добре, якщо швидкість не важлива, наприклад, коли утворюється емаль та дентин. Однак при ремонті рани оптимально, щоб рана заживала скрізь відразу, а не з країв. Тому тіло часто спочатку відкладає якусь форму будівельних лісів, наприклад, паршу. У стоматології штучні риштування можуть бути створені, щоб допомогти організму зцілитися, грунтуючись на наших знаннях про функції наземної речовини. Приклади штучних будівельних лісів включають деякі кісткові пластмасові матеріали, пародонтальні мембрани та матеріали. На додаток до їх структурної ролі як риштування, що направляє клітини в нові місця, молекули наземної речовини забезпечують клітини інформацією. Ця інформація розповідає осередкам, де вони розташовані, і що вони повинні робити. Наприклад, коли стовбурова клітина зв'язується з фібронектином, фібронектин може доручити стовбуровій клітині експресувати різні гени та диференціюватися в новий тип клітин, наприклад, одонтобласт, і почати секретувати дентин. Отримати клітини в правильне розташування приємно, але вони повинні знати, що робити, коли вони туди потрапляють. Це відбувається під час формування зуба і у відповідь на травму зуба. Коли ми дізнаємося більше про те, як подрібнена речовина навчає стовбурові клітини, ми покращуємо допомогу зубам у відновленні себе. Прикро, що багато підручників замазують ґрунтову речовину як просто желатиновий матеріал поза клітиною, саме тому ми зайшли в неї більш детально, а замість цього глянули над мітохондріями.
Волокна
Три позаклітинні білки були помітні під світловим мікроскопом століття тому, і вони були згруповані разом як волокна позаклітинного матриксу. Як і фібронектин та інші білки наземної речовини, волокна виділяються клітинами, званими фібробластами.
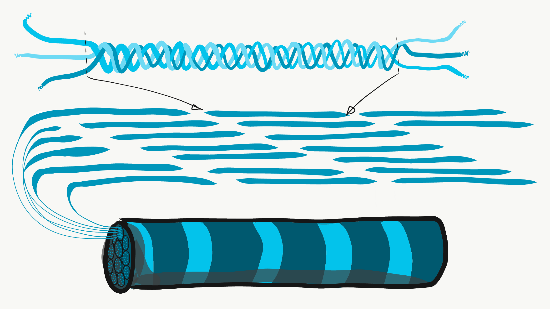
Колаген є найсильнішим з трьох волокон, і тому його часто називають структурним білком - він надає багатьом органам свою форму. Це волокно виготовлене з 3 спіральних α-спіралей, які, в свою чергу, згорнуті та зшиті разом, утворюючи дуже сильну макромолекулу з тією ж основною формою, що і мотузка. Вона дуже сильна, якщо тягнути на неї з кінців, але згинається, якщо докласти силу збоку. Наприклад, область прогресуючого карієсу на зубі може бути такою ж м'якою, як вологі корчі, оскільки вона містить свої колагенові волокна без великої кількості мінералів, які використовували для їх оточення. Колагенові волокна знаходяться в областях ротової порожнини, де важлива здатність протистояти силі, наприклад, в емалі або пародонтальної зв'язці. Насправді, він міститься в організмі людини, на нього припадає 25% вмісту нашого білка. Окрім того, що має структурну роль, колаген також діє як ешафот, подібний до фібронектину та ГК, що дозволяє клітинам мігрувати через довгі волокна. Отримання колагену з ReR (де він перекладається) в організм Гольджі для модифікації, і, нарешті, виділяється з клітини, вимагає надзвичайно великих бульбашок для транспортування. Ретикулярні волокна, які не показані на малюнку 1.11, виглядають як дрібна, схожа на павутинну сітку під мікроскопом. Пізніше було виявлено, що ретикулярні волокна - це різна форма колагену, але вони як і раніше називаються власною назвою, і часто отримують рівне з колагеном в списку ЕСУД волокон. Ця веб-подібна мережа білків не така сильна, але забезпечує достатньо рамки для клітин крові, щоб відпочити у всіх органах, таких як селезінка та лімфатичні вузли, на відміну від занурення на дно. Еластичні волокна тонше колагенових волокон, і часто виглядають як тонкі волоски в ЕСУД під мікроскопом. Вони виготовляються з іншого типу білка (еластину). Як випливає з назви, ці волокна можна розтягнути, а потім пружинити назад до початкової довжини. Це не те, що може зробити колаген (якщо ви не можете покласти долоні на підлогу, зберігаючи ноги прямими, колагенові волокна підколінного сухожилля та литкових м'язів та їх сухожилля обмежують діапазон руху. Регулярне розтягування може активувати активність фібробластів і подовжувати їх). Еластичні волокна виявляються у більшій кількості в областях ротової порожнини, які змінюють форму під час мови або ковтання, таких як м'яке піднебіння.
Ні в, ні поза:
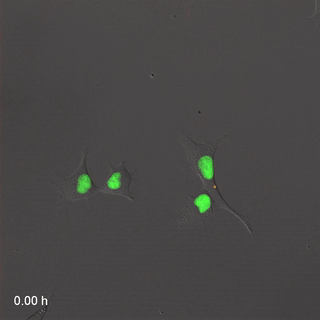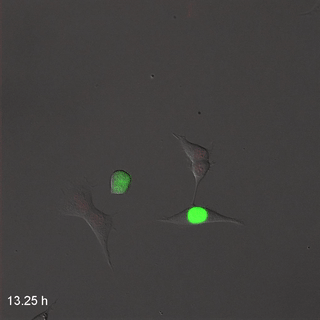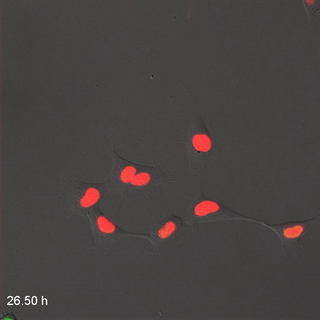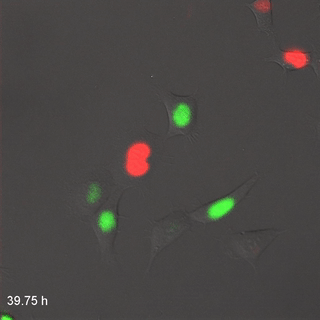
Мітоз
Розподіл клітин, або мітоз, процес, за допомогою якого одна клітина робить копію себе, виробляючи дві однакові дочірні клітини. На початку розвитку, коли ми зростаємо від однієї клітини до трильйона клітин, виникає багато мітозу. Коли клітина не зазнає мітозу, вона, як кажуть, знаходиться в інтерфазі. Це час, коли клітина може виконувати свою роботу, наприклад, виробляти волокна для нового ECM, або клітина може готуватися до мітозу. Перш ніж поділ клітин може відбутися, клітина повинна мати приблизно подвійну частину всього. Під час мітозу все ділиться навпіл між двома новими дочірніми клітинами. Не всі клітини здатні до мітозу - насправді більшість клітин у дорослої людини диференційовані і виконують завдання, вони занадто зайняті для розмноження. Ми говоримо, що ці невиліковно диференційовані клітини вийшли з клітинного циклу. Для відновлення пошкоджень тканини мають стовбурові клітини. Стовбурові клітини - це недиференційовані клітини, які здатні ділитися і диференціюватися на один або кілька різних типів клітин. Стовбурова клітина ділиться на дві дочірні клітини, одна дочка зазвичай залишається стовбуровою клітиною, а інша диференціюється на будь-який тип клітин, яким вона доручена. Таким чином, тканина може мати постійний запас стовбурових клітин, якщо стовбурові клітини не гинуть до того, як вони пройдуть мітоз. Коли ми стаємо старше, наші тканини не заживають так само, оскільки ми втратили стовбурові клітини. Взагалі кажучи, коли стовбурова клітина гине, її немає, інша стовбурова клітина не піддається мітозу, щоб виробити дві стовбурові клітини та замінити їх. Стовбурові клітини називаються виходячи з того, скільки різних типів клітин вони потенційно можуть стати. Однопотужні стовбурові клітини ротового епітелію стають кератиноцитами, і тільки кератиноцитами. Мультипотентні нейро-мезенхімальні стовбурові клітини перетворюються на дентин, пульпу, цемент і пародонтальну зв'язку. Всемогутня запліднена яйцеклітина стає кожною клітиною людини, плюс більше.

Щоб пройти інтерфазу і підготуватися до чергового раунду мітозу, клітини проходять через ряд контрольних точок клітинного циклу. Це реакції, що каталізуються ферментами, які перешкоджають прогресуванню до наступної фази до певної кількості продукту. Це встановлює базову швидкість поділу клітин, що допомагає забезпечити правильну кількість росту тканин. Проходження через контрольні точки клітинного циклу регулюється фосфорилюванням білків, званих циклінами. Цикліни - це фактори транскрипції, які активують гени, що дозволяють прогресувати через контрольну точку. Швидкість цього процесу може бути прискорена або сповільнена зовнішніми сигналами, такими як фактори росту. Фактори росту - це гормони, які виділяються в грунтову речовину тканини. Щільність і липкість подрібненої речовини впливає на те, наскільки сильно розсіюється фактор росту. Якщо дифузія обмежена, фактор росту лише прискорює зростання в локалізованій області. Це важливо при формуванні нових органів, таких як зуби. Інші фактори росту можуть поширюватися на широкій площі, особливо якщо вони виділяються в кров. Секреція гормону росту в крові, наприклад, дозволяє різні органи рости приблизно з однаковою швидкістю. Мутація в гені для фактора росту, або рецептора фактора росту, може призвести до посилення функції. Якщо клітини отримують постійний сигнал включення для проходження через клітинний цикл в результаті, ми називаємо ці гени онкогенами. Для отримання функції потрібна лише одна мутована копія гена рецептора. Цикліни та інші гени, які регулюють клітинний цикл, однак, називаються генами-супресорами пухлини. Щоб клітина втратила здатність регулювати контрольні точки клітинного циклу, обидві копії гена супресора пухлини повинні бути мутовані. У вашому автомобілі, це займе одну ногу, щоб наступити на газ занадто важко, але вам доведеться не вистачати обох ніг, щоб не мати змоги вдарити гальма. Зазвичай рак утворюється, коли одна клітина набуває мутації як онкогенів, так і до пар алелей генів супресора пухлини. Алель - це одна з двох копій гена, знайденого в одному положенні на двох гомологічних хромосомах. Навіть якщо це відбувається, є ще один шар захисту покритий наступний розділ.

Апоптоз
Всі клітини містять групу клітинно-поверхневих рецепторних білків і внутрішньоклітинних ферментів, які дозволяють їм піддаватися запрограмованої загибелі клітин, або апоптозу, коли вказано інструкції. Запрограмована загибель клітин має вирішальне значення для багатоклітинного життя, що є дивною справою. Без нього наші тіла були б сповнені дуже старих, що не функціонують клітин. Або ще гірше, оскільки клітини досягли кінця свого життя - що для епітеліальних і клітин крові дійсно короткий - вони вивільняють вміст своїх лізосом і мітохондрій. Як ви пам'ятаєте, вміст цих органел є сильнокислим, які пошкоджують або вбивають сусідні клітини. Якщо ці сусідні клітини загинули в результаті, вони теж вивільняли б свій лізосомальний і мітохондріальний вміст, завдаючи ще більшої шкоди. Коли це відбувається в організмі людини, це називається некрозом тканин. Це ще не все, мертва клітина викидає ДНК, а ДНК дійсно довга і тягуча і, як правило, дуже липка. Це може захопити інші клітини і запобігти їх міграції належним чином. Це чудово, якщо здорово, якщо клітина захоплює і вбиває бактерії і не проти померти в процесі, але не щось хороше, щоб зробити з сусідніми людськими клітинами. Тому клітинам доручають пройти апоптоз, коли вони досягають кінця свого життя, якщо імунна система визначила їх інфікованими або раковими, або просто більше не потрібні. Апоптоз гарантує, що перед тим, як клітина загине, вона нейтралізує рН своїх лізосом і мітохондрій і подрібнює її ДНК на безпечні дрібні шматочки. Під час розробки виробляється більше клітин, ніж потрібно, а зайві клітини видаляються пізніше організовано - подібно до того, як будівництво великої будівлі передбачає спочатку будівництво будівельних лісів, а ліси видаляються ближче до кінця проекту. Процес відновлення рани також передбачає надмірне виробництво клітин з подальшим їх організованим видаленням. Сподіваємось, це здасться логічним до того часу, коли ви закінчите цей курс. Під час відновлення рани включаються інструкції ДНК, які використовувалися клітинами, коли вони розвивалися ембріонально. Якщо ви хочете звучати фантазії, і чи не всі ми, ви можете сказати, що загоєння ран повторює ембріональний розвиток. Рекапітулювати означає знову заявити (повторити). Процес апоптозу починається або з внутрішнього, або зовнішнього сигналу. Наприклад, коли ДНК клітини стає занадто мутованою, або якщо під час мітозу є непарна кількість хромосом, це викликає апоптоз. Крім того, клітина може бути доручена пройти апоптоз з позаклітинного сигналу під назвою Фактор некрозу пухлини (ФНП). Серія ферментів, званих каспазами, активується, що в кінцевому підсумку призводить до нейтралізації кислот, руйнування ДНК і змушує клітину вибухати на численні дрібні шматочки, які можуть бути очищені макрофагами.
З'єднання між клітиною і клітиною
| Види клітинних з'єднань | Приклади |
|---|---|
| Клітинка-клітинка | Десмосоми, щільні стики, зазорні з'єднання |
| Клітина до ECM | Гемі-десмосоми |
Таблиця 1.2: Основні типи з'єднань клітин. З'єднання - це спеціалізовані групи білків на поверхні клітини або поблизу неї, які роблять з'єднання з якоюсь іншою структурою. Ці з'єднання можуть бути з іншими осередками, або до ECM, як зазначено в таблиці 1.2.

Десмосоми
Анкерні з'єднання (десмосоми) являють собою міцні зв'язки між двома клітинами. Анкерні з'єднання з'єднуються і закріплюють цитоскелет однієї клітини до її сусіда. Велика група клітин, закріплених між собою цими переходами, набагато сильніша як група. «Одна гілочка ламається, але пучок гілочок міцний» —Текумсе. Гемі-десмосоми - це половина десмосоми, закріпленої на ECM, наприклад, ущільнення між епітелієм ясен та неклітинною поверхнею зуба. Одним з багатьох білків в десмосомі є інтегран. Цей білок розпізнає і зв'язується з білками в ECM, такими як фібронектин. Коли білок інтегран клітини з'єднується з фібронектином, це не тільки закріплює цитоскелет клітини до ECM і закріплює клітину на місці, інтегран також сигналізує всередину клітини, дозволяючи ядру знати, в якому типі тканини знаходиться клітина. Перш ніж клітина може перейти на нове місце, вона повинна спочатку видалити її прив'язуючі з'єднання. Під час розвитку клітини мігрують на нові локації і утворюють нові структури. Під час загоєння ран стовбурові клітини відділяються від сусідів, мігрують в травмовану область і починають ділення клітин, щоб створити достатню кількість клітин для фіксації травми.

щільні місця з'єднання
Тугі з'єднання - це менші переходи між осередками. Щільні з'єднання повністю оточують клітину і створюють водонепроникне ущільнення між цією коміркою та іншою клітиною. Це служить для створення бар'єрів між однією частиною тіла та іншою, дозволяючи клітинам регулювати те, що проходить, а що ні. Це також надає клітинам апікально-базолатеральну полярність (або різницю між верхом і низом), що особливо важливо для епітелію. Верхівкова сторона епітеліальної клітини звернена до просвіту (порожнистого центру) органу, тоді як базолатеральная сторона найближча до нижньої сполучної тканини. Білки, синтезовані на ReR, можуть бути відправлені або в верхівковий, або базолатеральний ділянку плазматичної мембрани. Потрапивши туди, транс-мембранні білки верхівкової сторони клітини не можуть дифузіровать до плазматичної мембрани на базолатеральній стороні, оскільки кільце щільних з'єднань блокує їх рух.
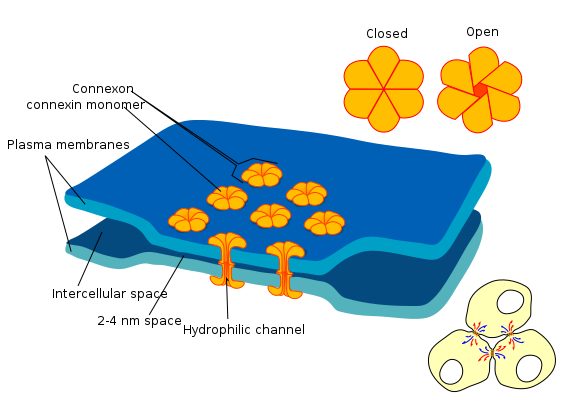
Розрив стиків
Розривні з'єднання (або коннексони) - це група білків, які утворюють невеликий прохід між клітинами, і можуть бути відкриті або закриті. Цей розрив дозволяє клітинам спілкуватися безпосередньо один з одним. Через те, як епітеліальні клітини з'єднані один з одним— в sheet— це спілкування відбувається через площину. Це один із способів того, як клітини знають своє положення щодо однієї з осей тіла, і це процес, який називається Planar Cell Polarity (PCP) (більше читання можна знайти тут). PCP - це полярність збоку в бік, тоді як апікально-базолатеральна полярність - зверху вниз. PCP дозволяє клітинам знати, в який бік вони спрямовані в тілі, гарантуючи, що структури, які вони утворюють, знаходяться не тільки в правильному місці, але і в правильній орієнтації. Наприклад, цей тип сигналізації дозволяє зубам формуватися так, що мовна сторона зуба звернена до мовної сторони, не звернена до щічної, мезіальної або дистальної сторін. Планарна клітинна полярність також допомагає зубам прорізуватися прямо в ротову порожнину, а не під кутом.
Глава 0* Глава 2