6.4: Скорочення м'язів
- Page ID
- 72842
Цілі навчання
- Опишіть кроки, що беруть участь у скороченні м'язів
Модель розсувної нитки стиснення
Щоб м'язова клітина скоротилася, саркомер повинен скорочуватися. Однак товсті і тонкі нитки - компоненти саркомери - не вкорочують. Натомість вони ковзають один за одного, внаслідок чого саркомер скорочується, тоді як нитки залишаються однакової довжини. Теорія ковзної нитки скорочення м'язів була розроблена з урахуванням відмінностей, що спостерігаються в названих смугах на саркомере при різних ступенях скорочення і розслаблення м'язів. Механізм скорочення полягає в зв'язуванні міозину з актином, утворюючи перехресні мости, які генерують рух нитки (рис. 6.7).
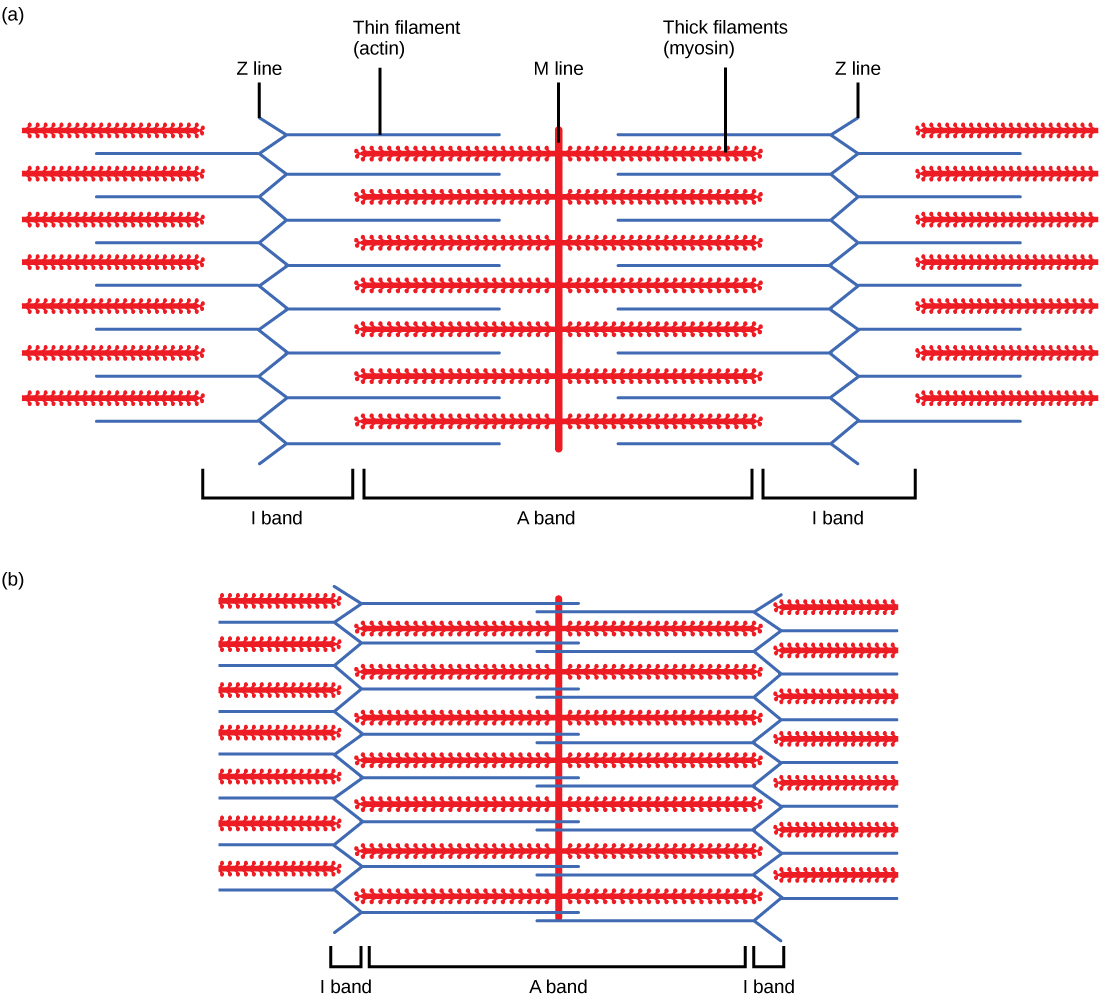
Малюнок 6.7. Коли (а) саркомер (b) стискається, лінії Z рухаються ближче один до одного, а група I стає меншою. Смуга А залишається однаковою шириною, і при повному стисненні тонкі нитки перекриваються.
Коли саркомер скорочується, деякі регіони скорочуються, тоді як інші залишаються однаковою довжиною. Саркомер визначається як відстань між двома послідовними Z дисками або лініями Z; коли м'яз скорочується, відстань між дисками Z зменшується. Зона Н - центральна область зони А - містить лише товсті нитки і скорочується під час скорочення. I смуга містить тільки тонкі нитки і також вкорочується. Смуга А не скорочується - вона залишається однаковою довжиною - але смуги різних саркомерів рухаються ближче один до одного під час скорочення, зрештою зникаючи. Тонкі нитки витягуються товстими нитками до центру саркомера, поки диски Z не наближаються до товстих ниток. Зона перекриття, в якій тонкі нитки і товсті нитки займають однакову площу, збільшується в міру просування тонких ниток всередину.
АТФ і скорочення м'язів
Рух скорочення м'язів відбувається, коли головки міозину зв'язуються з актином і тягнуть актин всередину. Для цієї дії потрібна енергія, яка забезпечується АТФ. Міозин зв'язується з актином в місці зв'язування на кулястому білку актину. Міозин має ще одне місце зв'язування АТФ, при якому ферментативна активність гідролізує АТФ до АДФ, виділяючи неорганічну молекулу фосфату і енергію.
АТФ зв'язування змушує міозин вивільняти актин, дозволяючи актину і міозину відділятися один від одного. Після того, як це станеться, знову пов'язаний АТФ перетворюється в АДФ і неорганічний фосфат Pi. Фермент в місці зв'язування на міозині називається АТФазою. Енергія, що виділяється при гідролізі АТФ, змінює кут головки міозину в «зведене» положення. Головка міозину потім знаходиться в положенні для подальшого руху, володіючи потенційною енергією, але АДФ і Пі все ще прикріплюються. Якщо сайти зв'язування актину покриті та недоступні, міозин залишатиметься у конфігурації високої енергії з гідролізованим АТФ, але все ще прикріпленим.
Якщо ділянки зв'язування актину розкриті, утворюється перехресний міст; тобто головка міозину охоплює відстань між молекулами актину та міозину. Потім Pi звільняється, дозволяючи міозину витрачати накопичену енергію як конформаційну зміну. Головка міозину рухається до лінії М, витягуючи актин разом з нею. У міру витягування актину нитки рухаються приблизно на 10 нм до лінії М. Цей рух називається силовим ударом, так як це крок, на якому виробляється сила. Коли актин тягнеться до лінії М, саркомер скорочується, а м'яз скорочується.
Коли головка міозину «зведена», вона містить енергію і знаходиться в високоенергетичній конфігурації. Ця енергія витрачається, коли головка міозину рухається через силовий удар; в кінці силового удару головка міозину знаходиться в низькоенергетичному положенні. Після силового удару виділяється АДФ; однак сформований поперечний міст все ще знаходиться на місці, а актин і міозин пов'язані між собою. АТФ потім може приєднуватися до міозину, що дозволяє циклу крос-моста почати знову і подальше скорочення м'язів може відбутися (рис. 6.8).
Перегляньте це відео, що пояснює, як сигналізується скорочення м'язів.
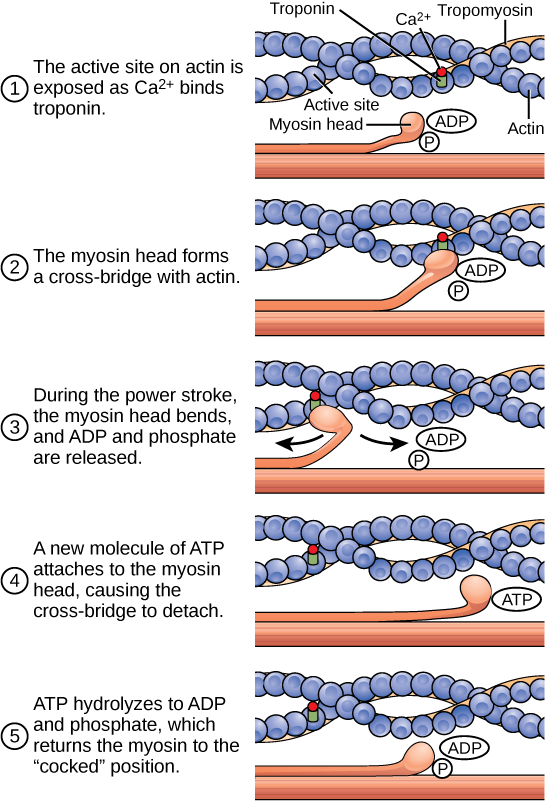
Малюнок 6.8. Показано цикл скорочення м'язів перехресного моста, який запускається зв'язуванням Ca2+ з активною ділянкою актину. З кожним циклом скорочення актин переміщається щодо міозину.
Вправа\(\PageIndex{1}\)
Яке з наведених нижче тверджень про скорочення м'язів вірно?
- Потужність удару відбувається, коли АТФ гідролізується до АДФ і фосфату.
- Силовий удар виникає, коли АДФ і фосфат дисоціюють з голівкою міозину.
- Силовий удар виникає при дисоціації АДФ і фосфату від активної ділянки актину.
- Потужність удару відбувається, коли Ca2+ пов'язує головку кальцію.
Перегляньте цю відеоанімацію скорочення м'язів перехресного моста.
регуляторні білки
Коли м'яз знаходиться в стані спокою, актин і міозин відокремлюються. Щоб актин не зв'язувався з активною ділянкою на міозині, регуляторні білки блокують місця молекулярного зв'язування. Тропоміозин блокує місця зв'язування міозину на молекулах актину, запобігаючи утворенню перехресних мостів і запобігаючи скороченню в м'язі без нервового введення. Тропонін зв'язується з тропоміозином і допомагає розташувати його на молекулі актину; він також пов'язує іони кальцію.
Щоб забезпечити скорочення м'язів, тропоміозин повинен змінити конформацію, розкриваючи міозінзв'язуючий ділянку на молекулі актину і дозволяючи формувати поперечний міст. Це може статися тільки в присутності кальцію, який зберігається в екстремально низьких концентраціях в саркоплазмі. Якщо вони присутні, іони кальцію зв'язуються з тропонином, викликаючи конформаційні зміни тропоніну, які дозволяють тропоміозину відійти від місць зв'язування міозину на актині. Після видалення тропоміозина між актином і міозином може утворитися перехресний міст, викликаючи скорочення. Перехресний мостовий цикл триває до тих пір, поки іони Ca2+ та АТФ більше не будуть доступні, а тропоміозин знову покриє місця зв'язування на актині.
Збудження-стиснення муфти
Збудження-скорочення зв'язку є ланкою (трансдукцією) між потенціалом дії, що генерується в сарколемі, і початком скорочення м'язів. Тригером для виділення кальцію з саркоплазматичної сітки в саркоплазму є нейронний сигнал. Кожне волокно скелетних м'язів контролюється руховим нейроном, який проводить сигнали від головного або спинного мозку до м'яза. Область сарколеми на м'язовому волокні, яка взаємодіє з нейроном, називається моторної торцевої пластиною. Кінець аксона нейрона називається синаптичним терміналом, і він фактично не контактує з торцевою пластиною двигуна. Невеликий простір, який називається синаптичною щілиною, відокремлює синаптичний термінал від торцевої пластини двигуна. Електричні сигнали рухаються по аксону нейрона, який розгалужується через м'яз і з'єднується з окремими м'язовими волокнами на нервово-м'язовому з'єднанні.
Здатність клітин спілкуватися електрично вимагає, щоб клітини витрачали енергію для створення електричного градієнта через свої клітинні мембрани. Цей градієнт заряду переноситься іонами, які диференційно розподілені по мембрані. Кожен іон надає електричний вплив і вплив концентрації. Подібно до того, як молоко з часом змішується з кавою без необхідності перемішувати, іони також розподіляються рівномірно, якщо їм дозволено це зробити. При цьому їм не дозволяється повертатися в рівномірно змішаний стан.
Натрій-калієва АТФаза використовує клітинну енергію для переміщення іонів K+всередині клітини та іонів Na+ назовні. Тільки це накопичує невеликий електричний заряд, але великий градієнт концентрації. У клітині багато K + і багато Na+ поза коміркою. Калій здатний виходити з клітини через K+ канали, які відкриті 90% часу, і це робить. Однак Na+ канали рідко відкриті, тому Na+залишається поза коміркою. Коли K+ залишає клітину, підкоряючись її градієнту концентрації, це фактично залишає негативний заряд позаду. Отже, у спокої існує великий градієнт концентрації для проникнення Na+ в клітину, і в клітці залишається накопичення негативних зарядів. Це потенціал спокою мембрани. Потенціал в цьому контексті означає поділ електричного заряду, який здатний виконувати роботу. Вимірюється вона в вольтах, точно так само, як і акумулятор. Однак трансмембранний потенціал значно менше (0,07 В), тому мале значення виражається мілівольтами (мВ) або 70 мВ. Оскільки внутрішня частина осередку негативна порівняно із зовнішньою, знак мінус означає перевищення негативних зарядів всередині комірки, −70 мВ.
Якщо подія змінить проникність мембрани до іонів Na+, вони потраплять в клітину. Що призведе до зміни напруги. Це електрична подія, яка називається потенціалом дії, яку можна використовувати як сигнал стільникового зв'язку. Зв'язок відбувається між нервами і м'язами через нейромедіатори. Потенціали дії нейрона викликають вивільнення нейромедіаторів з синаптичного терміналу в синаптичну щілину, де вони можуть потім дифузувати через синаптичну щілину і зв'язуватися з молекулою рецептора на моторній торцевій пластині. Моторна торцева пластина має стикові складки-складки в сарколемі, які створюють велику площу поверхні для нейромедіатора, щоб зв'язуватися з рецепторами. Рецептори насправді є натрієвими каналами, які відкриваються, щоб дозволити прохід Na+в клітину, коли вони отримують сигнал нейромедіатора.
Ацетилхолін (ACH) - це нейромедіатор, що виділяється руховими нейронами, який зв'язується з рецепторами в моторній торцевій пластині. Вивільнення нейромедіатора відбувається, коли потенціал дії рухається вниз по аксону рухового нейрона, що призводить до зміни проникності синаптичної термінальної мембрани і припливу кальцію. Іони Ca2+ дозволяють синаптичним везикулам рухатися і зв'язуватися з пресинаптичною мембраною (на нейроні) і вивільняти нейромедіатор з везикул в синаптичну щілину. Після звільнення синаптичним терміналом, АЧ дифундує через синаптичну щілину до торцевої пластини двигуна, де він зв'язується з рецепторами ACH. Як нейромедіатор зв'язується, ці іонні канали відкриваються, і іони Na+ перетинають мембрану в м'язову клітину. Це зменшує різницю напруг між внутрішньою і зовнішньою частиною осередку, що називається деполяризацією. Оскільки ACH зв'язується на торцевій пластині двигуна, ця деполяризація називається потенціалом кінцевої пластини. Потім деполяризація поширюється вздовж сарколеми, створюючи потенціал дії, оскільки натрієві канали, що прилягають до початкового місця деполяризації, виявляють зміну напруги та відкривають. Потенціал дії переміщається по всій клітині, створюючи хвилю деполяризації.
АЧ розщеплюється ферментом ацетилхолінестерази (AchE) на ацетил і холін. AchE знаходиться в синаптичній щілині, руйнуючи АЧ так, що він не залишається пов'язаним з рецепторами Ach, що призведе до небажаного розширеного скорочення м'язів (рис. 6.9).

Малюнок 6.9. Ця діаграма показує збудження-скорочення зв'язку в скороченні скелетних м'язів. Саркоплазматичний ретикулум - це спеціалізований ендоплазматичний ретикулум, який міститься в м'язових клітині.
Вправа\(\PageIndex{1}\)
Смертельний нервовий газ Зарин необоротно пригнічує ацетилхолінестеразу. Який вплив матиме зарин на скорочення м'язів?
Після деполяризації мембрана повертається в стан спокою. Це називається реполяризацією, під час якої закриваються напругою натрієві канали. Калієві канали тривають при 90% провідності. Оскільки плазматична мембрана натрій—калієва АТфаза завжди транспортує іони, стан спокою (негативно заряджений всередині відносно зовні) відновлюється. Період, безпосередньо наступний за передачею імпульсу в нерві або м'язі, в якому нейрон або м'язова клітина відновлює свою здатність передавати інший імпульс, називається рефрактерним періодом. Протягом рефрактерного періоду мембрана не може генерувати інший потенціал дії. Період вогнетривкості дозволяє чутливим до напруги іонним каналам повернутися до своїх конфігурацій спокою. Натрій-калієва ATPase постійно переміщує Na+назад з клітини, а K+назад у клітку, і K+витікає, залишаючи негативний заряд позаду. Дуже швидко мембрана реполяризується, так що вона знову може деполяризуватися.
Контроль м'язового напруження
Нейронний контроль ініціює утворення перехресних мостів актин-міозину, що призводить до вкорочення саркомеру, що бере участь у скороченні м'язів. Ці скорочення поширюються від м'язового волокна через сполучну тканину, щоб тягнути на кістки, викликаючи скелетний рух. Тяга, що чиниться м'язом, називається напругою, і величина сили, створюваної цим напругою, може варіюватися. Це дає можливість одним і тим же м'язам переміщати дуже легкі предмети і дуже важкі предмети. В окремих м'язових волокні кількість виробленого напруги залежить від площі поперечного перерізу м'язового волокна і частоти нервової стимуляції.
Кількість перехресних мостів, що утворюються між актином і міозином, визначає величину напруги, яку може виробляти м'язове волокно. Поперечні мости можуть утворюватися тільки там, де товсті і тонкі нитки перекриваються, дозволяючи міозину зв'язуватися з актином. Якщо утворюється більше перехресних мостів, більше міозину буде тягнути актин, і буде вироблятися більше напруги.
Ідеальна довжина саркомера при виробництві максимального натягу виникає, коли товсті і тонкі нитки перекриваються найбільшою мірою. Якщо саркомер в спокої розтягнутий повз ідеальну довжину спокою, товсті і тонкі нитки не перекриваються в найбільшій мірі, і може утворитися менше перехресних містків. Це призводить до меншої кількості міозинних голів, що тягнуть актин, і виробляється менша напруга. У міру вкорочення саркомеру зона перекриття зменшується, оскільки тонкі нитки досягають зони Н, яка складається з міозінних хвостів. Оскільки саме головки міозину утворюють перехресні мости, актин не зв'язується з міозином у цій зоні, зменшуючи напругу, що виробляється цим міофібром. Якщо саркомер укорочений, ще більше, тонкі нитки починають перекриватися один з одним, зменшуючи утворення поперечного моста ще більше і виробляючи ще менше натягу. І навпаки, якщо саркомер розтягнутий до точки, в якій товсті і тонкі нитки взагалі не перекриваються, не утворюються перехресні містки і не виробляється натяг. Така кількість розтягування зазвичай не відбувається, оскільки допоміжні білки, внутрішні сенсорні нерви та сполучна тканина протистоять екстремальному розтягуванню.
Первинною змінною, що визначає вироблення сили, є кількість міофібер всередині м'язи, які отримують потенціал дії від нейрона, який контролює це волокно. При використанні біцепса для підчеплення олівця рухова кора мозку сигналізує лише про кілька нейронів біцепса, і реагують лише кілька міофібер. У хребетних кожен міофібер повністю реагує, якщо стимулюється. Підбираючи піаніно, рухова кора сигналізує про всі нейрони в біцепсі, і кожен міофібер бере участь. Це близьке до максимальної сили, яку може виробляти м'яз. Як вже говорилося вище, збільшення частоти потенціалів дії (кількість сигналів в секунду) може збільшити силу трохи більше, тому що тропоміозин залитий кальцієм.
Вправа\(\PageIndex{1}\)
Навчіть свого однолітку про події під час скорочення м'язів, від надходження нервового сигналу до генерації руху, що живиться м'язом. Коли ви закінчите, запитайте свого однолітку, які терміни або кроки ви пропустили або не пояснили добре. Нехай ваш одноліток заповнить прогалини. Якщо не було прогалин, ваш однолітк може кинути вам виклик з деякими питаннями щодо вашого пояснення. Пам'ятайте, що один із способів перевірити, чи навчаєтесь ви, - це мати можливість передавати свої знання іншій людині.