10.3: Скорочення та розслаблення м'язових волокон
- Page ID
- 1198

Цілі навчання
- Опишіть компоненти, що беруть участь у скороченні м'язів
- Поясніть, як м'язи скорочуються і розслабляються
- Опишіть модель ковзної нитки скорочення м'язів
Послідовність подій, які призводять до скорочення окремого м'язового волокна, починається з сигналу - нейромедіатора, АЧ-від рухового нейрона, що іннервує це волокно. Місцева мембрана волокна деполяризується, оскільки позитивно заряджені іони натрію (Na +) потрапляють, викликаючи потенціал дії, який поширюється на решту мембрани, деполяризуватиметься, включаючи Т-канальці. Це запускає вивільнення іонів кальцію (Ca ++) із зберігання в саркоплазматичному ретикулумі (SR). Потім Ca ++ ініціює скорочення, яке підтримується АТФ (рис.\(\PageIndex{1}\)). Поки іони Ca ++ залишаються в саркоплазмі, щоб зв'язуватися з тропонином, який тримає сайти, що зв'язують актин, «неекранованими», і до тих пір, поки АТФ доступний для їзди на велосипеді крос-мосту та витягування актинових ниток міозином, м'язове волокно буде продовжувати скорочуватися до анатомічної межі.
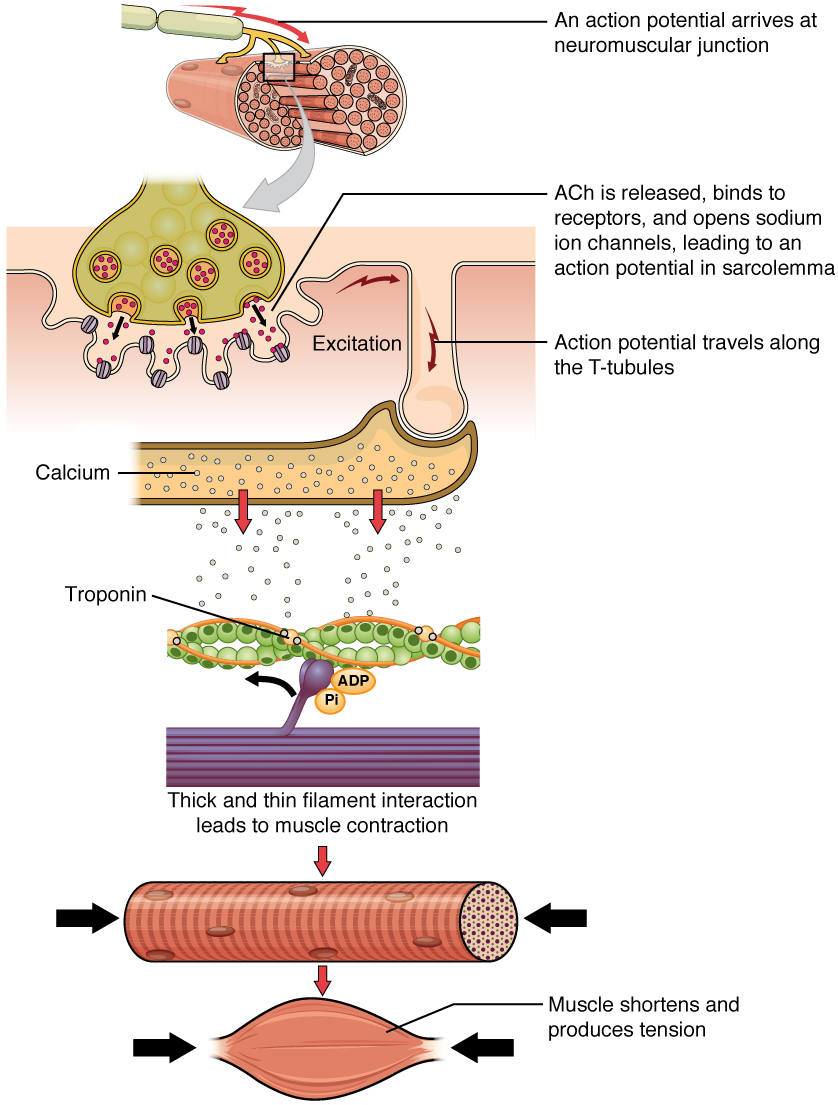
Скорочення м'язів зазвичай припиняється, коли закінчується сигналізація від рухового нейрона, який реполяризує сарколему і Т-канальці, і закриває в СР кальцієві канали з напругою. Потім іони Ca ++ перекачуються назад в SR, що призводить до того, що тропоміозин відновлює (або повторно покриває) місця зв'язування на актинових нитках. М'яз також може перестати скорочуватися, коли вона закінчується АТФ і стає втомленою (рис.\(\PageIndex{2}\)).


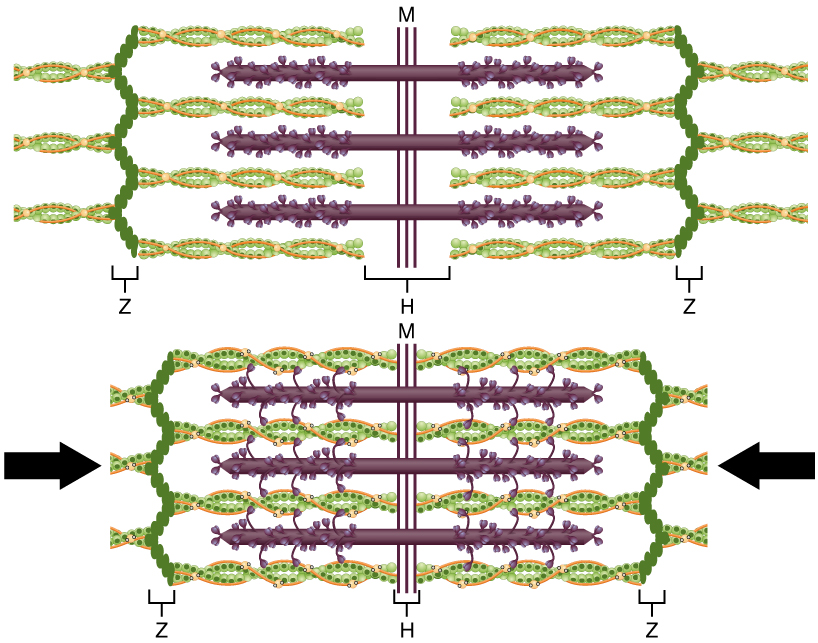
Молекулярні події скорочення м'язових волокон відбуваються всередині саркомерів волокна (див. Рис.\(\PageIndex{3}\)). Скорочення поперечно-смугастого м'язового волокна відбувається, коли саркомери, лінійно розташовані всередині міофібрил, скорочуються, коли головки міозину тягнуть актинові нитки.
Область, де перетинаються товсті і тонкі нитки, має щільний вигляд, так як між нитками мало місця. Ця зона, де перетинаються тонкі і товсті нитки, дуже важлива для скорочення м'язів, оскільки це місце, де починається рух ниток. Тонкі нитки, закріплені на своїх кінцях Z-дисками, не поширюються повністю в центральну область, яка містить лише товсті нитки, закріплені біля їх основи на місці, званому M-лінією. Міофібрила складається з багатьох саркомерів, що проходять по його довжині; таким чином, міофібрили та м'язові клітини скорочуються, коли саркомери скорочуються.
Модель ковзної нитки стиснення
Коли сигналізує руховий нейрон, волокно скелетних м'язів стискається, коли тонкі нитки витягуються, а потім ковзають повз товстих ниток всередині саркомерів волокна. Цей процес відомий як модель ковзної нитки скорочення м'язів (рис.\(\PageIndex{3}\)). Ковзання може відбуватися лише тоді, коли міозинзв'язуючі ділянки на актинових нитках піддаються ряду кроків, які починаються з входження Са ++ в саркоплазму.
Тропоміозин - це білок, який намотується навколо ланцюгів актинової нитки і покриває міозінзв'язуючі ділянки, щоб запобігти зв'язуванню актину з міозином. Тропоміозин зв'язується з тропонином, утворюючи комплекс тропонін-тропоміозин. Комплекс тропонін-тропоміозин перешкоджає зв'язуванню міозінних «головок» з активними ділянками на актинових мікрофіламентах. Тропонін також має місце зв'язування іонів Са ++.
Щоб ініціювати скорочення м'язів, тропоміозин повинен оголити міозинзв'язуючий ділянку на актинової нитці, щоб забезпечити утворення поперечного моста між мікрофіламентами актину та міозину. Перший крок у процесі скорочення полягає в тому, щоб Ca ++ зв'язувався з тропонином, щоб тропоміозин міг ковзати від місць зв'язування на актинових пасмах. Це дозволяє голівкам міозину зв'язуватися з цими відкритими місцями зв'язування та утворювати перехресні мости. Потім тонкі нитки витягуються головками міозину, щоб ковзати повз товстих ниток до центру саркомеру. Але кожна голова може витягнути лише дуже коротку відстань, перш ніж вона досягне своєї межі, і її потрібно «повторно звести», перш ніж вона зможе знову потягнути, крок, який вимагає АТФ.
АТФ і скорочення м'язів
Для того, щоб тонкі нитки продовжували ковзати повз товстих ниток під час скорочення м'язів, головки міозину повинні витягувати актин у місцях зв'язування, від'єднуватися, повторно півень, прикріплюватися до більшої кількості місць зв'язування, тягнути, від'єднати, повторно півень тощо Цей повторний рух відомий як цикл перехресного моста. Цей рух міозінних голів схожий на весла, коли окремі ряди човен: Весло весла (головки міозину) тягнуть, піднімаються з води (від'єднуються), переставляються (повторно зведені), а потім знову занурюються, щоб тягнути (рис.\(\PageIndex{4}\)). Кожен цикл вимагає енергії, а дія міозінних головок в саркомерах, що повторюються натягують на тонкі нитки, також вимагає енергії, яка забезпечується АТФ.
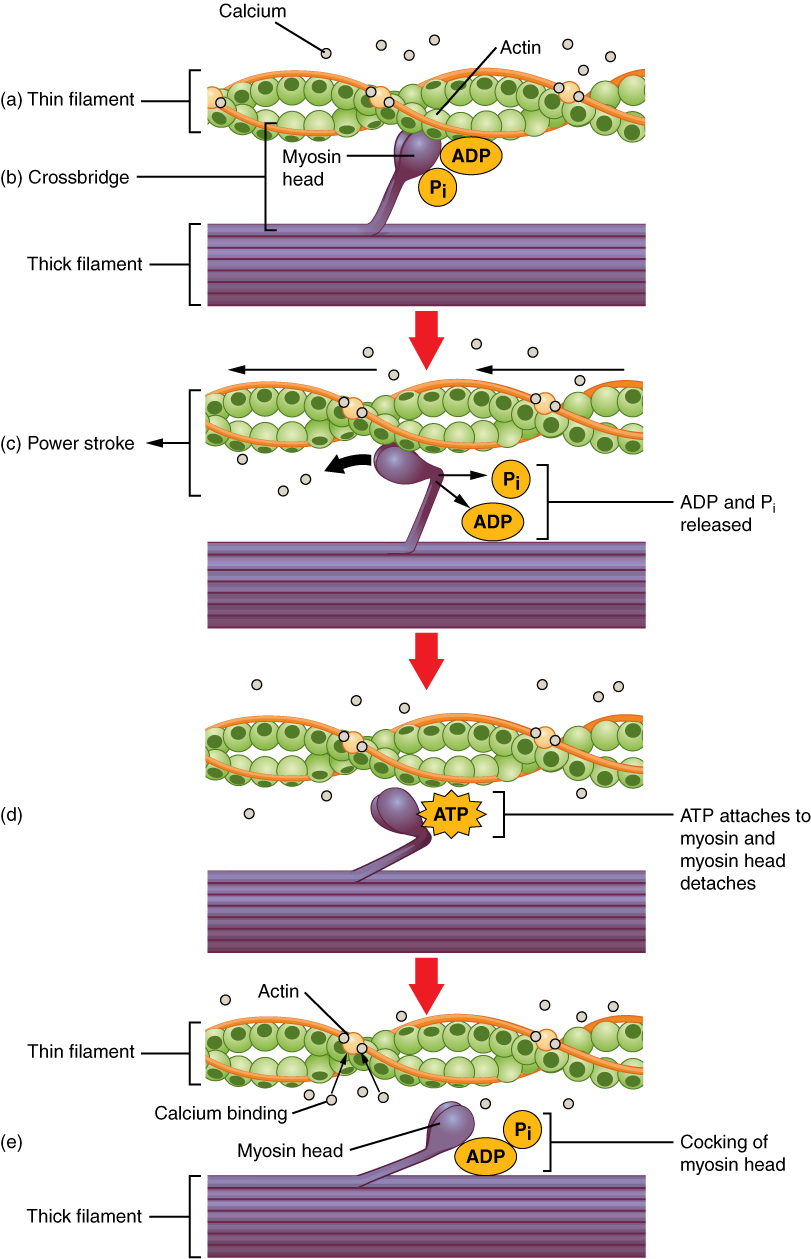
Формування перехресного моста відбувається, коли головка міозину прикріплюється до актину, тоді як аденозиндифосфат (ADP) та неорганічний фосфат (P i) все ще пов'язані з міозином (рис. 10.3.4.a, b.). P i потім звільняється, в результаті чого міозин утворює більш сильне прикріплення до актину, після чого головка міозину рухається до М-лінії, витягуючи актин разом з нею. Коли актин витягується, нитки рухаються приблизно на 10 нм у напрямку до М-лінії. Цей рух називається силовим ходом, так як рух тонкої нитки відбувається на цьому кроці (рис\(\PageIndex{4}\). в.). При відсутності АТФ головка міозину не відірветься від актину.
Одна частина головки міозину прикріплюється до місця зв'язування на актині, але головка має інше місце зв'язування для АТФ. АТФ-зв'язування змушує головку міозину відриватися від дії (рис\(\PageIndex{4}\). д.). Після цього АТФ перетворюється в АДФ і Р i за рахунок внутрішньої АТФАЗной активності міозину. Енергія, що виділяється при гідролізі АТФ, змінює кут головки міозину в зведене положення (рис\(\PageIndex{4}\). е.). Головка міозину тепер знаходиться в положенні для подальшого руху.
При зведенні головки міозину міозин знаходиться в високоенергетичної конфігурації. Ця енергія витрачається, коли головка міозину рухається через силовий удар, а в кінці силового удару головка міозину знаходиться в низькоенергетичному положенні. Після силового удару виділяється АДФ; однак сформований поперечний міст все ще знаходиться на місці, а актин і міозин пов'язані між собою. Поки АТФ доступний, він легко прикріплюється до міозину, цикл крос-моста може повторюватися, а скорочення м'язів може тривати.
Зверніть увагу, що кожна товста нитка приблизно 300 молекул міозину має множинні головки міозину, і багато перехресних мостів утворюються і розриваються безперервно під час скорочення м'язів. Помножте це на всі саркомери в одній міофібрилі, всі міофібрили в одному м'язовому волокні, і всі м'язові волокна в одному скелетному м'язі, і ви можете зрозуміти, чому так багато енергії (АТФ) потрібно, щоб підтримувати роботу скелетних м'язів. Насправді, саме втрата АТФ призводить до суворості смертності, що спостерігається незабаром після того, як хтось помирає. Якщо подальше виробництво АТФ не можливе, немає АТФ, доступного для міозінних голів, щоб від'єднатися від ділянок, що зв'язують актин, тому перехресні мости залишаються на місці, викликаючи жорсткість скелетних м'язів.
джерела СПС
АТФ постачає енергію для скорочення м'язів. На додаток до своєї прямої ролі в циклі крос-моста, АТФ також забезпечує енергією для активно-транспортних насосів Ca ++ в СР. Скорочення м'язів не відбувається без достатньої кількості АТФ. Кількість АТФ, що зберігається в м'язах, дуже низька, достатньо лише для живлення скорочень на кілька секунд. Оскільки він руйнується, АТФ слід швидко регенерувати та замінювати, щоб забезпечити стійке скорочення. Існує три механізми, за допомогою яких АТФ може регенеруватися: обмін креатинфосфату, анаеробний гліколіз, ферментація і аеробне дихання.
Креатинфосфат - це молекула, яка може зберігати енергію в своїх фосфатних зв'язках. У м'язі, що відпочиває, надлишок АТФ передає свою енергію креатину, виробляючи АДФ і креатинфосфат. Це діє як запас енергії, який можна використовувати для швидкого створення більшої кількості АТФ. Коли м'яз починає скорочуватися і потребує енергії, креатинфосфат передає свій фосфат назад в АДФ, утворюючи АТФ і креатин. Ця реакція каталізується ферментом креатинкінази і відбувається дуже швидко; таким чином, АТФ, отриманий з креатинфосфату, забезпечує перші кілька секунд скорочення м'язів. Однак креатинфосфат може забезпечити лише приблизно 15 секунд енергії, в цей момент потрібно використовувати інше джерело енергії (рис.\(\PageIndex{5}\)).

Оскільки АТФ, що виробляється креатинфосфатом, виснажується, м'язи звертаються до гліколізу як джерела АТФ. Гліколіз - це анаеробний (не залежний від кисню) процес, який розщеплює глюкозу (цукор) для отримання АТФ; однак гліколіз не може генерувати АТФ так швидко, як креатинфосфат. Таким чином, перехід на гліколіз призводить до більш повільної швидкості доступності АТФ до м'яза. Цукор, який використовується при гліколізі, може забезпечуватися глюкозою в крові або шляхом метаболізму глікогену, який зберігається в м'язі. Розпад однієї молекули глюкози виробляє дві АТФ і дві молекули піровиноградної кислоти, які можуть використовуватися при аеробному диханні або при низькому рівні кисню, перетвореному в молочну кислоту (\(\PageIndex{5}\)рис. б).
При наявності кисню піровиноградну кислоту використовують при аеробному диханні. Однак, якщо кисень недоступний, піровиноградна кислота перетворюється в молочну кислоту, що може сприяти втомі м'язів. Це перетворення дозволяє переробляти фермент NAD + з NADH, який необхідний для продовження гліколізу. Це відбувається під час напружених фізичних вправ, коли потрібна велика кількість енергії, але кисень не може бути достатньо доставлений до м'язів. Сам гліколіз не може бути витриманий дуже довго (приблизно 1 хвилина м'язової активності), але він корисний для полегшення коротких сплесків високої інтенсивності виходу. Це пояснюється тим, що гліколіз не дуже ефективно використовує глюкозу, виробляючи чистий приріст двох АТФ на молекулу глюкози та кінцевий продукт молочної кислоти, яка може сприяти втомі м'язів у міру її накопичення.
Аеробне дихання - це розпад глюкози або інших поживних речовин у присутності кисню (O 2) з утворенням вуглекислого газу, води та АТФ. Приблизно 95 відсотків АТФ, необхідного для відпочинку або помірно активних м'язів, забезпечується аеробним диханням, яке відбувається в мітохондріях. Входи для аеробного дихання включають глюкозу, що циркулює в крові, піровиноградну кислоту та жирні кислоти. Аеробне дихання набагато ефективніше анаеробного гліколізу, виробляючи приблизно 36 АТФ на молекулу глюкози проти чотирьох від гліколізу. Однак аеробне дихання не може бути витримано без стійкого надходження O 2 до скелетної мускулатури і відбувається набагато повільніше (\(\PageIndex{5}\)рис. в). Щоб компенсувати, м'язи зберігають невелику кількість надлишкового кисню в білках називають міоглобіном, що дозволяє більш ефективно скорочувати м'язи і менше втоми. Аеробні тренування також підвищують ефективність кровоносної системи, так що O 2 може подаватися до м'язів протягом більш тривалих періодів часу.
М'язова втома виникає, коли м'яз більше не може скорочуватися у відповідь на сигнали нервової системи. Точні причини м'язової втоми до кінця не відомі, хоча певні фактори корелювали зі зменшенням м'язового скорочення, яке відбувається під час втоми. АТФ необхідний для нормального скорочення м'язів, а так як резерви АТФ зменшуються, функція м'язів може знижуватися. Це може бути більше фактором у короткому, інтенсивному виході м'язів, а не стійких, менших зусиль інтенсивності. Накопичення молочної кислоти може знизити внутрішньоклітинний рН, впливаючи на активність ферментів і білків. Дисбаланси в рівнях Na + і K + в результаті деполяризації мембран можуть порушити відтік Ca ++ з SR. Тривалі періоди тривалих фізичних вправ можуть пошкодити SR та сарколему, що призведе до порушення регулювання Ca ++.
Інтенсивна м'язова активність призводить до кисневого боргу, який є кількістю кисню, необхідного для компенсації АТФ, що виробляється без кисню під час скорочення м'язів. Кисень необхідний для відновлення рівнів АТФ і креатинфосфатів, перетворення молочної кислоти в піровиноградну кислоту, а в печінці - для перетворення молочної кислоти в глюкозу або глікоген. Інші системи, що використовуються під час фізичних вправ, також вимагають кисню, і всі ці комбіновані процеси призводять до збільшення частоти дихання, що виникає після фізичних навантажень. Поки не буде виконана киснева заборгованість, споживання кисню підвищується, навіть після припинення фізичних навантажень.
Розслаблення скелетних м'язів
Розслаблення волокон скелетних м'язів, і, в кінцевому рахунку, скелетних м'язів, починається з рухового нейрона, який перестає випускати свій хімічний сигнал, АЧ, в синапс на NMJ. М'язове волокно буде реполяризуватися, що закриває ворота в SR, де випускався Ca ++. Насоси, керовані АТФ, переміщають Ca ++ з саркоплазми назад в SR. Це призводить до «переекранування» актинових зв'язуючих ділянок на тонких нитках. Без можливості утворювати перехресні мости між тонкими і товстими нитками м'язове волокно втрачає напругу і розслабляється.
Сила м'язів
Кількість скелетних м'язових волокон в даній м'язі генетично визначається і не змінюється. М'язова сила безпосередньо пов'язана з кількістю міофібрил і саркомерів всередині кожного волокна. Фактори, такі як гормони та стрес (і штучні анаболічні стероїди), що діють на м'язи, можуть збільшити вироблення саркомерів і міофібрил всередині м'язових волокон, зміна називається гіпертрофією, що призводить до збільшення маси і основної маси в скелетних м'язах. Так само зменшення використання скелетних м'язів призводить до атрофії, де кількість саркомерів і міофібрил зникає (але не кількість м'язових волокон). Зазвичай кінцівка в гіпсі показує атрофовані м'язи при знятті гіпсу, а деякі захворювання, такі як поліомієліт, показують атрофовані м'язи.
РОЗЛАДИ...
М'язова система
М'язова дистрофія Дюшенна (МДД) - прогресуюче ослаблення скелетних м'язів. Це одне з декількох захворювань, які в сукупності називають «м'язовою дистрофією». МДД викликається нестачею білка дистрофіну, який допомагає тонким ниткам міофібрил зв'язуватися з сарколемою. Без достатньої кількості дистрофіну м'язові скорочення викликають сарколему розриву, викликаючи приплив Ca ++, що призводить до пошкодження клітин та деградації м'язових волокон. Згодом, у міру накопичення м'язових пошкоджень, втрачається м'язова маса, розвиваються більші функціональні порушення.
МДД - це успадковане розлад, викликане аномальною Х-хромосомою. Вона в першу чергу вражає чоловіків, і зазвичай діагностується в ранньому дитинстві. DMD зазвичай спочатку з'являється як труднощі з рівновагою і рухом, а потім прогресує до неможливості ходити. Він продовжує прогресувати вгору в тілі від нижніх кінцівок до верхньої частини тіла, де впливає на м'язи, відповідальні за дихання та кровообіг. Зрештою, це спричиняє смерть через дихальну недостатність, і ті, хто страждає, як правило, не живуть за 20 років.
Оскільки DMD викликаний мутацією в гені, який кодує дистрофін, вважалося, що введення здорових міобластів пацієнтам може бути ефективним лікуванням. Міобласти - це ембріональні клітини, відповідальні за розвиток м'язів, і в ідеалі вони несуть здорові гени, які могли б виробляти дистрофін, необхідний для нормального скорочення м'язів. Цей підхід був значною мірою невдалим у людей. Недавній підхід передбачав спробу збільшити виробництво м'язів утропіну, білка, подібного до дистрофіну, який може бути в змозі взяти на себе роль дистрофіну та запобігти клітинному пошкодженню.
Огляд глави
Саркомер - це найменша скоротлива частина м'язи. Міофібрили складаються з товстих і тонких ниток. Товсті нитки складаються з білка міозину; тонкі нитки складаються з білка актину. Тропонін і тропоміозин є регуляторними білками.
Скорочення м'язів описується ковзною ниткою моделі скорочення. ACH - це нейромедіатор, який зв'язується на нервово-м'язовому з'єднанні (NMJ), щоб викликати деполяризацію, і потенціал дії рухається вздовж сарколеми, щоб викликати виділення кальцію з SR. Ділянки актину піддаються впливу після того, як Са ++ потрапляє в саркоплазму зі свого сховища SR, щоб активувати комплекс тропонін-тропоміозин так, що тропоміозин зміщується від ділянок. За поперечним перемиканням міпозінових головок стикуються в актинові вузли, що зв'язують, слідує «силовий хід» - ковзання тонких ниток товстими нитками. Харчування ударів здійснюється від АТФ. Зрештою, саркомери, міофібрили та м'язові волокна скорочуються, щоб виробляти рух.
Інтерактивні запитання щодо посилань
Вивільнення іонів кальцію ініціює м'язові скорочення. Перегляньте це відео, щоб дізнатися більше про роль кальцію. (а) Що таке «Т-трубочки» і яка їх роль? (b) Будь ласка, також опишіть, як актин-зв'язування сайтів доступні для перехресного зв'язування з міозиновими головками під час скорочення.
Відповідь: (а) Т-канальці є внутрішніми розширеннями сарколеми, які викликають вивільнення Ca ++ від SR під час потенціалу дії. (b) Ca ++ зв'язується з тропоміозином, і це відсуває стрижні тропоміозину від місць зв'язування.
Переглянути питання
Q. в розслабленому м'язі міозинзв'язуючий ділянку на актині блокується на ________.
Тітін А.
Б. тропонин
C. міоглобін
Д. тропоміозин
Відповідь: D
Q. Відповідно до моделі ковзної нитки, місця зв'язування на актині відкриваються при ________.
А. рівень креатину фосфату підвищується
B. Підвищення рівня АТФ
C. підвищення рівня ацетилхоліну
D. рівень іонів кальцію підвищується
Відповідь: D
В. Клітинна мембрана м'язового волокна називається ________.
А. міофібрил
Б. сарколема
С. саркоплазма
D. міофіламент
Відповідь: B
Q. розслаблення м'язів відбувається при ________.
А. іони кальцію активно транспортуються з саркоплазматичного ретикулума
Б. іони кальцію дифузійні з саркоплазматичного ретикулуму
С. іони кальцію активно транспортуються в саркоплазматичний ретикулум
D. іони кальцію дифузно в саркоплазматичний ретикулум
Відповідь: C
Q. під час скорочення м'язів поперечний міст відшаровується при ________.
А. головка міозину зв'язується з молекулою АДФ
Б. головка міозину зв'язується з молекулою АТФ
С. іони кальцію зв'язуються з тропонином
D. іони кальцію зв'язуються з актином
Відповідь: C
Q. Тонкі і товсті нитки організовані в функціональні одиниці, які називаються ________.
А. міофібрили
Б. міофіламенти
C. Т-трубочки
Д. саркомери
Відповідь: D
Питання критичного мислення
Питання: Як вплинули б м'язові скорочення, якщо волокна скелетних м'язів не мали Т-канальців?
А. без Т-канальців потенційна провідність дії у внутрішню частину клітини відбуватиметься набагато повільніше, викликаючи затримки між нервовою стимуляцією та скороченням м'язів, що призводить до більш повільних, слабких скорочень.
Питання: Що викликає поперечно-смугасте поява скелетної м'язової тканини?
А. темні смуги А і світлі I смуги повторюються вздовж міофібрил, а вирівнювання міофібрил в клітці змушують всю клітину здаватися смугастою.
Питання: Як вплинули б м'язові скорочення, якщо АТФ був повністю виснажений в м'язовому волокні?
А. без АТФ головки міозину не можуть від'єднатися від вузлів, що зв'язують актин. Всі «застряглі» перехресні мости призводять до жорсткості м'язів. У живої людини це може викликати такий стан, як «судоми письменника». У недавно померлої людини це призводить до суворості мортису.
Глосарій
- аеробне дихання
- вироблення АТФ в присутності кисню
- АТФАЗА
- фермент, який гідролізує АТФ до АДФ
- креатин фосфат
- фосфаген використовується для накопичення енергії від АТФ і передачі її в м'язи
- гліколіз
- анаеробний розпад глюкози до АТФ
- молочна кислота
- продукт анаеробного гліколізу
- кисневий борг
- кількість кисню, необхідного для компенсації АТФ, що виробляється без кисню під час скорочення м'язів
- потужність ходу
- дія міозину, що тягне актин всередину (до лінії М)
- піровиноградна кислота
- продукт гліколізу, який може бути використаний при аеробному диханні або перетворений в молочну кислоту